Social Echolocation calls - the case of Pipistrellus kuhlii
Pipistrellus kuhlii
Pipistrellus kuhlii (PIKU) is considered to be acoustically a typical Pipistrellus, with echolocation calls in open situations dropping to a very low slope. The ends of pulses can be very variable, often showing a kink upwards or a sudden drop downwards in frequency. The flattest calls may not show much of a downsweep at all. Some calls can show an overall rise, rather than decrease, in frequency and this can be seen in many genera of bats which normally produce very low slope calls when flying in the open.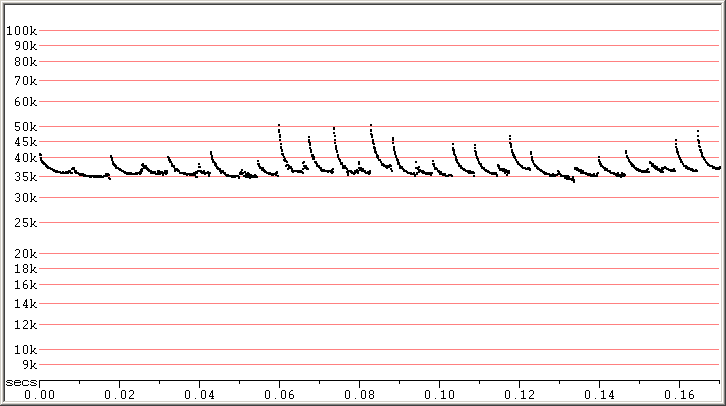
Fig 1. Solaris Resort, Croatia, 3 Sep 2014 showing a variety of search-phase calls. F7, compressed. Fc at lower end
of range for PIKU.
Like other Pipistrellus, as PIKU go into clutter, their calls get shorter in duration, with steeper Sc and a bit of an increase in Fc. Most calls in clutter still differ from typical calls of Myotis in having a marked reduction in slope ("hook") near the end of the call. Some individual pulses can closely resemble Myotis calls, but extended sequences usually won't, because these are calls of Pipistrellus in clutter where things change rapidly, while the similar calls of Myotis will be made in more open situations and show less variability.
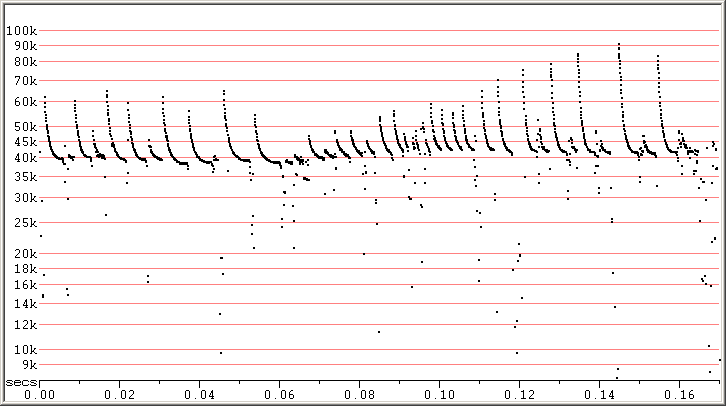
Fig 2. Zagreb, Croatia, 9 Sep 2010 showing transition from lower to high clutter in search phase. F7, compressed.
Note that none of these pulses looks much like a Myotis! All show the "hook" at the bottom to some degree.
This is all the same bat, and with Fc being at the higher end of the range for PIKU.
Good examples of PIKU going into clutter can be seen when the bats go into attack phase. Attack phase shows a transition from lower clutter search phase calls into the feeding buzz, and in doing so, it illustrates a broad range of call types which could also be seen in search phase sequences. Usually, as the bat goes into higher clutter, the frequency rises somewhat, though in Pipistrellus the rise in frequency is usually only a few kiloHerz.
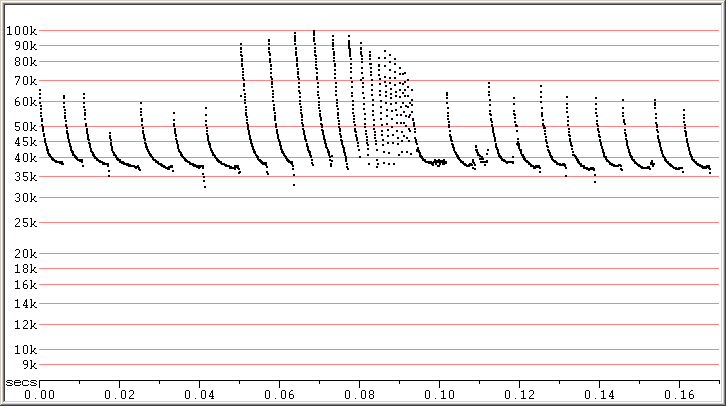
Fig 3. Zagreb, Croatia, 9 Sep 2010 showing search phase calls in some clutter and a feeding buzz. F7, compressed.
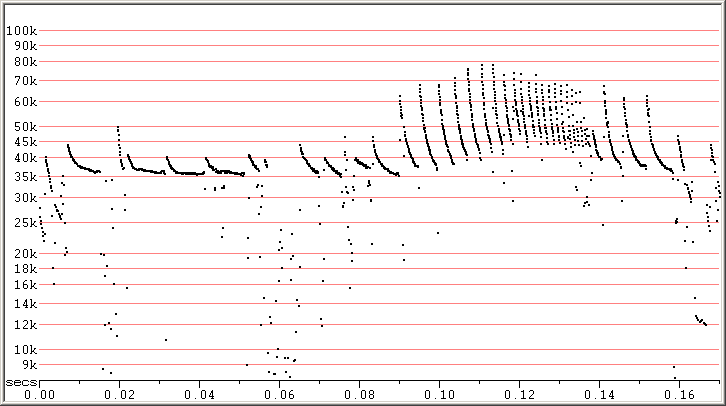
Fig 4. Zagreb, Croatia, 9 Sep 2010 showing a much more dramatic feeding buzz with a 10 kHz rise in frequency.
Note the rather Myotis like pulses in the attack phase. The songs at each end of the sequence, and the single steep
pulse above 0.02 are likely from a different bat. F7 compressed.
PIKU echolocation calls mostly have Fc in the 35 to 41 kHz range. They slightly overlap in frequency with P. pipistrellus (PIPI) but widely overlap with P. nathusii (PINA) to the point where it is unlikely that PINA and PIKU could reliably be distinguished by their echolocation calls.
Songs of PIKU are quite distinctive, being lower in frequency and typically longer in duration, despite fewer pulses, than for PIPI or P. pygmaeus (PIPY). They are quite different from the much more complex songs of PINA. Songs of all these Pipistrellus are well documented in Middleton, Froud and French (2014).
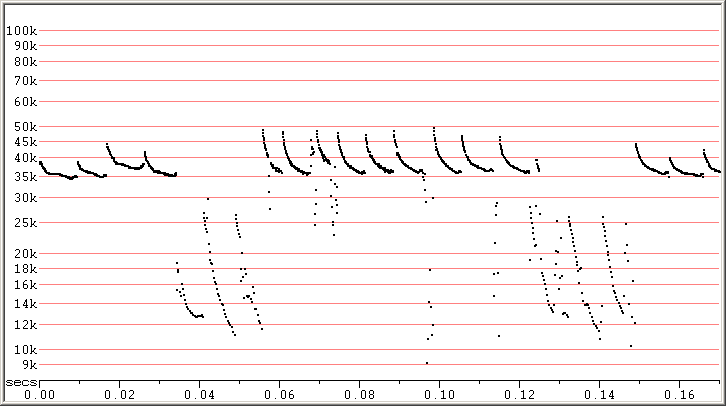
Fig 5. Zagreb, Croatia, 2 Sep 2010 showing two songs amongst normal echolocation calls.
What I found in Croatia
During the 2014 EBRS at Solaris Resort near Sibenik, Croatia, I made many recordings of bats using both passive and active recording. I was surprised to see many calls which were unfamiliar to me. These looked like Myotis in shape with Fc which could suggest Myotis myotis, but the total experience was all wrong for Myotis - too much variation for their low clutter calls. At first, I was thinking these might have been brief, high clutter sequences of Serotines or Leislers, but again the total picture was wrong - it makes no sense to keep seeing short, high clutter sequences of these species without seeing other types of calls. I noticed that these mystery calls always seemed closely associated with PIKU songs, but that didn't mean much, because PIKU songs were so common in the area.After the EBRS, I took part in the Capacity Building Workshop at Skradin, Croatia. One of the many joys of that place was sitting in the early morning hours watching bats from the porch of the guest house. It was very common to see the same types of calls I had encountered at Solaris, and I soon became confident these were made by PIKU. They were almost always associated with PIKU songs and I was able to watch bats giving these calls and in the same sequence, transitioning into normal PIKU echolocation calls. I also watched bats making these calls transition directly into normal attack and feeding buzz sequences. So it was evident that these calls were a different calling mode of PIKU, but were still being used as echolocation calls in the pursuit of insects.
For convenience, I will refer to them as Social Echolocation (SE) calls. I discuss the logic of that below.
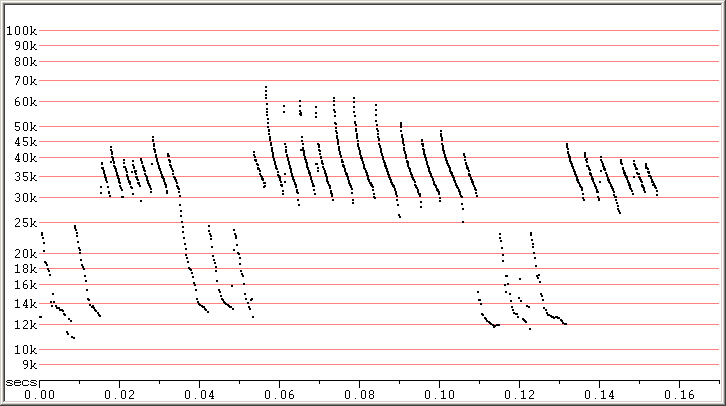
Fig 6. Skradin, Croatia, 5 Sep 2014 showing SE calls with Fc down to 30 kHz, and 3 songs. F7 compressed.
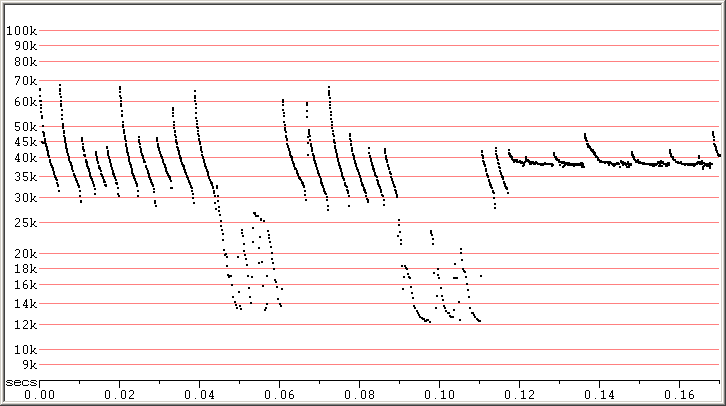
Fig 7. Skradin, Croatia, 5 Sep 2014 showing SE calls contrasted with normal calls. It is uncertain if all these calls
were from the same bat. F7 compressed.
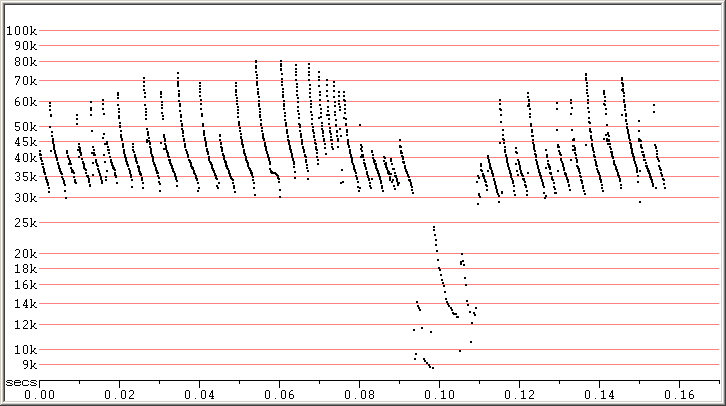
Fig 8. Skradin, Croatia, 5 Sep 2014 showing SE calls progressing into, then returning from, an attack phase,
followed by a song. F7 compressed.
Fig 9. Skradin, Croatia, 5 Sep 2014 showing SE calls transitioning to normal echolocation briefly, then back to
SE calls and two songs. This whole sequence is from one bat, though the songs may have been made by a different
bat. F7 compressed.
For those who want to see things in full-spectrum, here is a segment of a file made by Kate Barlow, showing two typical PIKU echolocation pulses, the second much fainter, and between them, two SE pulses from a second bat. Note the difference in frequency between the two SE pulses, with the first showing Fc about 30 kHz and the second with Fc above 35 kHz. This kind of variation is often in SE pulses of PIKU, but in this case, it is also notable that the second, higher pulse, is also showing a lower Sc than the previous pulse. This combination of higher Fc and lower Sc is not what would be expected in adjacent pulses from the same bat emitting either type of echolocation calls, and maybe a sign that the second pulse is part of a transition from SE to normal calls.
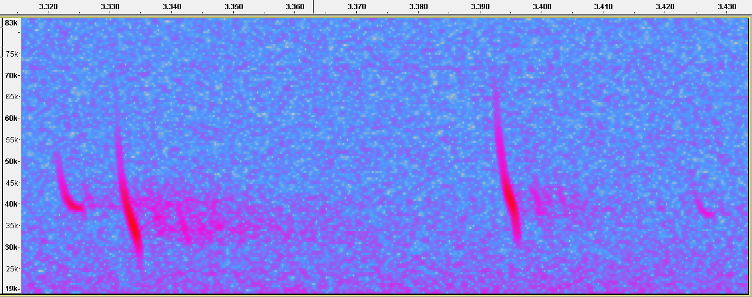
Fig 10. Skradin, Croatia, 10 Sep 2014. Full-spectrum recording made by Kate Barlow showing two SE calls between two normal calls.
Overall, SE calls cover a wide range of Fc values. While normal echolocation calls can show Fc from 35 to 41 kHz, the SE calls may be a lot lower, down to a bit below 30 kHz or up to above 40 kHz. The SE calls, like the normal echolocation calls, cover a wide rang of slopes depending on the clutter in which the Bat is flying, but even in low clutter they are much steeper than normal echolocation calls and overall droop downwards at the end like a Myotis. The terminal droop may result in Fmin even lower than 25 kHz.
So what are SE calls?
Most social calls we see from flying bats are brief compared to the time between echolocation calls. That makes sense, because if a bat is depending on its echolocation, it can't just stop echolocating and make other types of acoustic signals for an extended period. Such social calls are typically very different to the echolocation pulses, usually being a lot lower in frequency and often of completely different shape. The songs of Pipistrelles are good examples of this.On the other hand, some types of social calls are just echolocation calls with bits added on to them, and these may be produced over an extended time period. In this case, the individual pulses can still play the same roles as the echolocation calls, so the bat can echolocate while conveying social meaning. One European example of such calls might be Daubenton's Bat calls with the upswept hook at the start (type C from Middleton, Froud and French, 2014). Another might be the Plecotus calls which drop well below 20 kHz as a terminal downsweep after low slope echolocation calls. I am not sure where the boundary lies between social calls and low clutter echolocation calls in Plecotus, and this ambiguity is alluded to in Russ (2012) at the bottom of page 163. Elsewhere I have seen other cases, such as Corynorhinus townsendii, Eptesicus fuscus, Tadarida brasiliensis and several species of Myotis in north America.
SE calls seem to be a different case, where an echolocation function is retained in social calls, but the echolocation elements are quite different from those seen in normal echolocation calls. In PIKU, the SE calls seem closely associated with songs. When I first realised that PIKU was the species involved, I looked through my many recordings of PIKU giving songs but failed to find obvious SE calls amongst many of them, especially in France. However, having looked again at these recordings, I now think it may be the case that PIKU always echolocate with SE calls when they are singing. Determining this is not as simple as it may seem. There are several complicating factors:
- the songs are much lower in frequency than the echolocation calls, and can therefore be detected from much further away.
- there are often multiple bats present, and this seems especially so when PIKU are singing, compared to other Pipistrellus.
- PIKU live in places where there are other species, such as Myotis myotis, to which the SE calls might be attributed.
- The SE calls do superficially resemble some high clutter calls of PIKU and may be overlooked for that reason.
My assumption is that these SE calls are primarily social calls, being part of the whole song flight display, but they also have roles as echolocation calls, and as such show a typical range of call types depending on clutter. They are used to catch prey. They are produced in extended sequences containing only SE calls and often songs. They are rather variable compared to other PIKU echolocation calls and this suggests they are generated in situations of greater clutter. They are not part of the usual search phase continuum, though broadly overlapping with it in frequency. Essentially, compared to the normal PIKU echolcation calls, they show a separate continuum of search phase calls as clutter varies. They are lower in frequency, higher in Sc and without the typical Pipistrellus tendency to rapidly decrease in slope towards the end of the pulse. They also show a much greater tendency to droop downwards in frequency at the end of a pulse.
Are there SE calls in other Pipistrellus?
So far, I have not found any evidence of equivalent calls in PIPY, PIPI or PINA. In the cases of PIPY and PIPI, I have looked at many recordings where the only echolocation calls are typical Pipistrellus calls, not SE calls. In more complex situations, SE calls might be overlooked as coming from Myotis, and more species of Myotis might cause confusion with any SE calls produced by PIPY or PIPI than is the case with PIKU.The situation with PINA is more difficult, as I have far less experience with PINA and most if not all of the songs I have recorded have been produced from roost sites rather than in flight. PINA shouldn't be expected to make SE calls when not flying, although that is certainly not a necessity, as other bat species will echolocate from inside the entrances to roosts.
How to recognise SE calls
To recognise SE calls, it is almost essential to use compressed displays, because only those can show multiple pulses in sufficient detail to appreciate their shapes. As with any acoustic ID problem, a combination of compressed and truetime views is much more useful than either alone, since it not only reveals the shapes of pulses, but also their temporal relationships, which can resolve questions such as how many bats were recorded. Compressed and truetime views are complementary - each providing important data.Based on what I have seen in PIKU, I expect any SE calls from Pipistrellus to be lower in frequency (specifically Fc) than normal echolocation pulses given by the same individual, and to have a body (portion of least slope) which is steeper. If PIPI was making SE calls, they might reach down to below 40 kHz and look superficially like various species of Myotis. I would expect Myotis calls of similar shape to be in sequences showing greater regularity in pulse shape and call repetition rate, though this could be a difficult distinction if only a few pulses can be seen.
Ultimately, the best evidence of SE calls would come from observations of Bat calls morphing from one type to the other within the same sequence, but at least within PIKU, this was not commonly encountered.
Consequences
These observations considerably expand the range of frequencies which can be expected from PIKU echolocation calls, including all of the range of Hypsugo savii (HYSA) calls. However, the SE calls are not like HYSA calls except in sharing common Fc values, as they are much steeper and shorter in duration. SE calls of PIKU also expand the range of call shapes which can be expected from PIKU, to call types which much more closely resemble calls of Myotis than Pipistrellus. It may also be that SE calls of PIKU are unique to that species and might be a useful identification character in separating PIKU from PINA, although the value of this will be reduced by the fact that SE calls seem closely linked to songs of PIKU, which are very distinctive in themselves.Since these observations, I have seen suggestions of SE call types in other species (not Pipistrellus). In these cases there are no distinctive songs - rather the bats seem to be making social displays of some kind with the main components being echolocation calls which are different from their usual echolocation calls.
Comments invited!
I would greatly appreciate any comments on the above, positive, negative or anything else! The best way to contact me is at the Email address shown at my home page. It would be especially interesting to hear of people finding evidence of SE type calls in other species, and particularly in other Pipistrellus.References
Middleton, N., Froud, A. and French, K. (2014) Social Calls of the Bats of Britain and Ireland. Exeter: Pelagic Publishing.Russ, J. (2012) British Bat Calls - A Guide to Species Identification. Exeter: Pelagic Publishing.
Back to: Bat Notes contents, Anabat Contents, Hom