Back to: Bat Notes
contents, Anabat
Contents, Home
In December 2007, December 2008, and late November to early December 2009, I recorded bats in Sabah, Malaysia on the island of Borneo. Most time was spent at the Danum Valley Field Center (DVFC), but recordings were also made in the township of Kota Kinabalu and at the Kinabatangan River. The trips were organized by Simon Robson from James Cook University in Townville, as part of biology courses at the university. On each trip, some students ran projects involving the capture of bats, which provided an opportunity to get to know some of the species in the area and to associate recorded calls with known species. However, many of the species recorded acoustically were not captured, though the identities of some of those can be inferred from other information. This writing is an attempt to characterise the call types recorded there, whether or not the species are known.
I am indebted to Simon Robson for making my presence in Sabah possible, and for his help in the field. I would also like to thank both Jen Parsons and Tamara Inkster for the profound contributions they made in supervising the bat projects and maintaining the integrity of the data collected. Glenn Hoye joined us in 2008, and contributed valuable insights into species identification issues. And of course, the many students who took part contributed by their enthusiasm and hard work over long hours.
Currently only 20 of the 30 species recorded can be assigned to species, and some of those are in doubt because the species could not be reliably identified in the hand, or because identifications are tentative as the bats were never captured. Furthermore, some of the species captured and identified have very faint and/or very ambiguous calls which may not be much use for identifying those species under wild conditions. Even so, I don't have any doubt about the value of acoustics as a tool for surveying bats, and my hope is that what I have written below will help people put acoustic identification on a stronger footing. Published material on the calls of bats in the region have concentrated primarily on other aspects of acoustics, while my main interest is in the use of acoustics for species identification. The latter requires much more detailed accounts than have previously been presented, because bat calls vary so much and there is usually broad overlap in call types between two or more species.
What's below should be thought of as a work in progress - as a starting point from which improvements will gradually lead to refinements of identification criteria. I expect that several of the species unidentified below will be readily recognisable when more is known about bat calls in the region. Probably, several of them will be distinctive enough, once someone has the opportunity to connect the calls to the species producing them.
I would also like to point out that much can be learned from watching the bats, either in the twilight or using spotlights. Bat-watching is little practiced outside of Europe, primarily because the task seems so daunting to people without that sort of background (ie looking at birds which are hard to identify, such as distant hawks or dark shearwaters). While it is true that bat-watching is nearly always limited by the inability to use other methods to confirm an identity, it is still an important tool because the combination of acoustics and visual observations is often much more powerful than using either method by itself. This is true even when visual observations are made at a very basic level, but in time, I expect that the art of bat-watching will become more main-stream and more useful, just as field-identification criteria have evolved to something really powerful in the avian world.
I rely heavily in my field-work on the use of a portable system (Anabat) which allows me to see spectrograms of bat calls in real time on a device which I can carry around in one hand. I firmly believe that such devices will become totally mainstream for bat-watching, as they provide vastly better information than other approaches to acoustic characterisation. While this view may be contentious, I see little ultimate value in the use of systems like heterodyne or time-expansion. Newer systems providing full-spectrum displays in near realtime may be thought of as providing better information than the frequency division system used by Anabat. However, these displays impose their own costs, and cannot compete with frequency-division for rapid access and immediate association of call types to visually observed bats. Full-spectrum displays are also inherently blurry, and it is much more difficult to quickly see the features of most importance for species identification.
I want to illustrate this last point with the case of Emballonurides versus Molossides. It is certainly true that calls given by some species of each family can be easily confused - especially when they are flying high and straight. This is one case where full-spectrum methods make things a lot easier, because the spectral signatures of these two families are immediately and obviously different, in ways which aren't readily perceived using Anabat. However, I don't find these two groups particularly difficult to distinguish in the field. Most of the time, I can see immediately which family I am detecting, just from more subtle features of the way the call sequence progresses. It certainly wouldn't be worth my while to deal with the ponderously slow and blurry displays, heavy power consumption and bulk of full-spectrum equipment just to resolve those few cases where the ambiguity is greatest.
All calls were all recorded using an Anabat SD1 bat detector and displayed in real time on an IPAQ 2215 or 2495 PDA. Wherever possible, I attempted to see the bats producing the calls, either by watching them in the twilight or by illuminating them with a portable, 100 Watt spotlight powered by lightweight, 2 to 3 AH NiMH batteries. I recorded field notes on a small, digital voice-recorder, and later embedded these notes into the Anabat files produced or wrote notes into Word documents.
When bats were captured, I attempted to record their echolocation calls upon release. To accomplish this, I stood close to the person releasing the bats, and if the situation allowed it, I would try to follow the bat on foot after its release. Where possible, I avoided directly illuminating the released bat with the central beam of the spotlight, because this tends to cause the bat to flee the area. Instead, I would try to keep just enough light on the bat to see where it was and be confident that the calls recorded were from that bat. As the bat moved off into the distance, I would illuminate it more strongly, to try to keep it in sight as long as feasible.
Such hand-releases are an essential tool for associating echolocation calls with particular species, but they are only ever likely to result in recordings of a limited part of a species' repertoire for most species. The strength of this approach comes from seeing some of the calls a bat can generate, and then associating them with other calls produced in the wild by free-flying bats, where calls produced in open situations (low or zero clutter) are often produced along with higher clutter calls seen when a bat comes near objects or during attack-phase sequences. In this way, a picture can be built up of the range of call types a bat is capable of producing. Contrary to the impression often created in the literature, search-phase calls cover a very wide range of call types for most species, and a thorough knowledge of this variation is required in order to understand how species can be distinguished and where their calls can overlap.
For examination of recorded calls in the lab after a night of recording., I used AnalookW software (version 3.6a to 3.8q).
Most recordings were made while walking around on tracks and roads within 1 km of the Danum Valley Field Centre (DVFC), and from the open back of a truck on road transects extending up to 7.5 km ENE from the field centre. Some species were only recorded upon release after capture.
Calls were examined carefully and classified into categories according to their frequencies and shapes. The categories were given names reflecting both properties.
Screen dumps were used to make pictures of representative call sequences from the AnalookW windows, and these were then manipulated using Microsoft Paint to form composite diagrams illustrating the main features of a variety of call types from each putative species recognised.
1) A Field Guide to the Mammals of Borneo, Payne and Francis, 2007. (MOB)
2) Kingston, Jones, Akbar and Kunz, 2003. Alternation of Echolocation Calls in 5 Species of Aerial-feeding Insectivorous Bats from Malaysia. J. Mamm, 84(1), 205-215. (KJAK)
3) Kingston, Liat and Akbar, 2009. Bats of Krau Wildlife Reserve. Penerbit Universiti Kebangsaan Malaysia. (KLA)
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
30 categories of call types were recognised. 13 categories have been associated with species captured, identified in the hand and released by students at the Danum Valley Field Centre. A further 4 categories are associated with species captured and released, but where there is some doubt about the identity of the bats concerned.
Species where identification is considered uncontentious were:
Family Hipposideridae
Family Rhinolophidae
Family Nycteridae
Family Vespertilionidae
Species where there is a question of identity were:
Family Hipposideridae
Hipposideros ater (but possibly H. dyacorum)
Family Emballonuridae
Emballonura monticola (but possibly E. alecto)
Family Vespertilionidae
Kerivoula intermedius ( but possibly K. minuta)
Myotis muricola or M. ater or both
Visual observations, in combination with published call descriptions, suggest that some call types can be confidently assigned to Cheiromeles torquatus, Tadarida plicata and Saccolaimus saccolaimus, to bats of the genus Miniopterus or of the family Emballonuridae. Notes and illustrations appear in the Appendix.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
It is important to appreciate that this list of categories is very tentative. Further experience is likely to reveal that several categories are in fact composite - that more than one species produces calls which have been put together here into one category. It is also possible that some categories are artificial, in that the same bat produces calls classified here into more than one category.
The hope is that by producing a list of categories in this manner, clarification will result, and that this will be a step towards refining knowledge of these bats. In generating this list, many points were noticed that would have been lost without this exercise.
Many call types can be confused with others. A good example of this is the confusion that can exist between F27 and F23, despite the fact that these bats belong to different families and their acoustic behaviours are very different. But many of their calls look very similar.
Bats are best recorded when alone. When there are more than one bat present, they tend to interact and produce calls not so typical of their species. The same problem occurs around sources of food - although streetlights are widely seen as good places to encounter bats, they are actually bad places to record bat calls, because the bats are producing such a wide range of call types and they are influenced by the presence of other bats, food and inert structures. The most useful calls are recorded from a bat flying alone in the open.
Danum Valley is an area of often intense bat activity, and it was often difficult to record bats without others appearing in the same recording. This made interpretation of some of the call types more difficult than it would have been in a place with less activity. For the same reason, it was often difficult to be sure a bat being observed visually was the same individual being recorded acoustically.
Despite these caveats, the categories
recognised here are mostly distinctive enough to be readily identified
with a
little practice.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
In this section, notes on the features of the different call categories are presented, along with illustrations showing call features and variation.
These categories are generally ordered by frequency from highest to lowest, but only within broad call-type classes. Categories are named by a one character prefix, which denotes the shape of the call, followed by a number which denotes the Characteristic Frequency (Fc) of typical examples. Note that call types are named for the lowest clutter calls given by that species. Most species produce a wide range of call types depending on the degree of clutter (proximity to sound-reflecting objects) in which they are flying. Usually, but not always, calls given in the lowest clutter (sometimes called commuting calls) are the most distinctive call types.
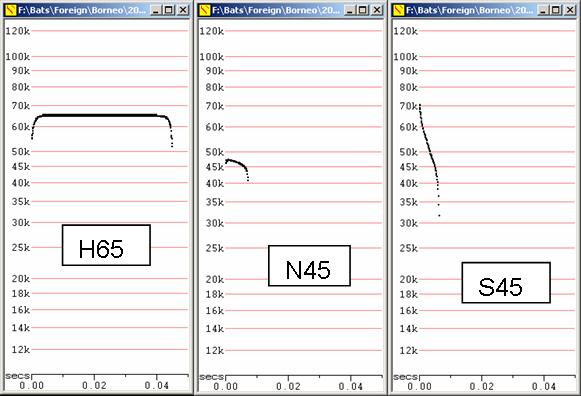
In some cases, the names attributed to call type were artificially changed to avoid duplication, For example, Hipposideros ridleyicalls and Rhinolophus creaghi could both be referred to as H65 type bats, but one is called H64 to avoid confusion, without any inference that there is a meaningful difference in frequency.
Call shape prefixes are:
H Very distinctive call types of Horseshoe Bats, (Rhinolophidae and Hipposideridae). Distinctive in being of high duty cycle with most of the call appearing at a constant or almost constant frequency, perhaps with an initial upsweep and terminal downsweep. Fc is also the Maximum Frequency (Fmax). In general, Rhinolophides give calls of longer duration, higher duty cycle and more constant frequency than Hipposiderides. Could potentially be confused with N.
N Call in which Fc is close to Fmax, like calls of Horseshoe Bats, but with a lower duty cycle and usually with a much less constant maximum frequency. Mostly dominated by the terminal downsweep, but many variations on this theme, so some potential confusion possible with H calls. Most energy is emitted on the second harmonic.
S Steep, generally linear calls, though may be much more complex, with two or three slope changes. Characterised by high slopes even in low clutter.
B Calls with a steep initial downsweep, followed by a fairly sudden reduction in slope near the bottom of the call. Such calls may lie somewhere between C and J type calls, or S and J type calls, and these categories may grade into each other.
J Calls which tend to flatten or hook back up after a pronounced initial downsweep, to form a reversed J shape.
C Curvilinear calls, in which the flattest calls (lowest slope) are of low slope but still noticeably curved. No clear distinction from F calls, and both may be followed by a brief droop in frequency in a terminal downsweep. A single species might produce both C and F type calls. The distinction is maintained here because it seems useful in distinguishing between categories, without any implication of taxonomic significance.
F Flat calls, though usually not with zero slope. Essentially linear calls, but maybe with a droop at the end, and of low slope. May also start with a very short downsweep. See under C above.
D Low slope calls with a pronounced droop in frequency at the end, occupying a significant part of the total call. May be actually higher clutter calls of a species which in lower clutter would give calls more appropriately labeled as C or F.
There are many variations on all these call types, but they are used here as a rough guide. Many calls will be encountered which are different from any of these, or which are borderline between two categories. The same bat may make a variety of call types within one sequence, but for the above purposes, broad categories believed to represent a single species, will be named after the flattest, lowest calls known from that species.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
For species where identification in the hand may not be straightforward, a section called Basis provides notes on what criteria were used to identify the animals recorded.
Taxonomic uncertainties, combined with the difficulty of in-the-hand identification for some species, which partly results from inadequate field guides, makes identification of many species tentative. Until better field identification criteria are published, the field biologist is constrained to make the best possible use of available information, but this is likely to often to mistakes. These may not be resolved if voucher specimens or genetic material were not collected, but there are many reasons why this is not always feasible. Apart from anything else, it is often pointless recording bats in an enclosed space, so it is best to release animals soon after capture and record them as they take flight. Since many bats usually produce atypical calls in such situations, or at best produce only a subset of their repertoire, the release of as many bats as possible enhances the probability of obtaining sequences of calls which are useful for subsequent recognition of free-flying animals.
A section called Observations gives information on what the bats looked like in free flight, and anything else about their behaviour or occurrence.
The bulk of each species account is dealing with its acoustic properties, under the heading Echolocation.
Pteropodidae
Emballonuridae
Megadermatidae
Nycteridae
Rhinolophidae
Hipposideridae
Vespertilionidae
Miniopteridae
Molossidae
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Category Sp if known
H155 probably Hipposideros ater (? dyacorum)
H150 Hipposideros sabanus
H115 Hipposideros cervinus
H87 Rhinolophus acuminatus
H82 Rhinolophus borneensis
H65 Hipposideros ridleyi
H64 Rhinolophus creaghi
H62 Hipposideros diadema
H60 Rhinolophus sedulus
H50 Rhinolophus trifoliatus
H38 Rhinolophus sp
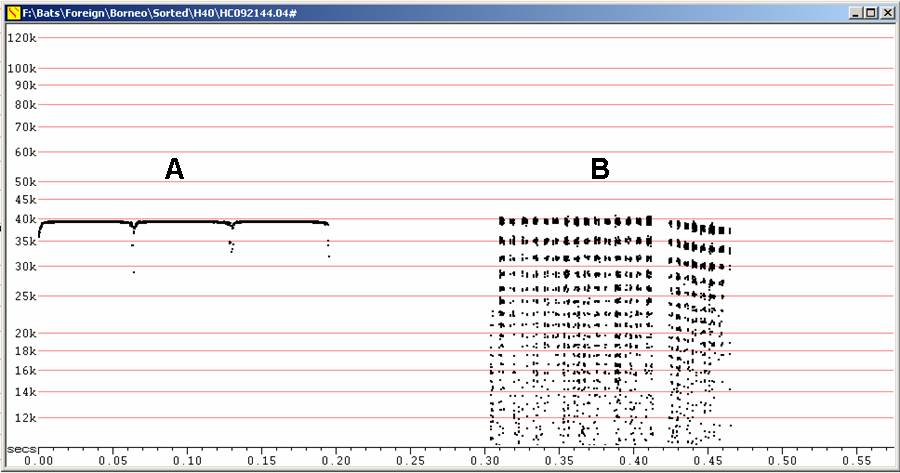
N45 Emballonura sp (monticola or alecto)
S140 Kerivoula hardwickii
S100 Kerivoula papillosa
S90 Kerivoula sp (intermedius or minuta)
C55 Miniopterus sp
S70 Nycteris javanica
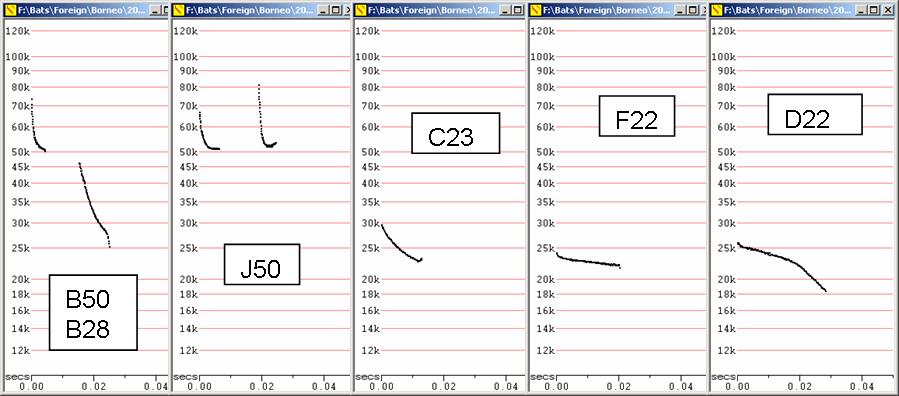
B50 Water-skimming Myotis
S50 Myotis, possibly M. horsfieldii
J50 Myotis prob muricola (? ater)
C40 Miniopterus sp
F23 Tadarida plicata
F27 Taphozous sp
F22 Saccolaimus saccolaimus
D22 Cheiromeles torquatus
C20 possibly Hesperoptenus tomesi
F16 probably Mops mops
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Some doubt about identity. One individual captured on 3 Dec 2008 seemed easily resolved to either Hipposideros ater or H. dyacorum. Appeared somewhat intermediate between these two, based on MOB. FA ca 41.5 mm. No lateral leaflets. Apparently adult - many scars on wings.
Body colour grey, without obvious contrast between dorsum and venter, venter certainly not white. Ears more like MOB depiction of dyacorum, markedly pointed and more triangular than rounded. Nose structure like MOB depiction of ater in morphology, but much more like depiction of dyacorum in colour. Colour greyish pink with posterior noseleaf darker. Internarial septum much more like depiction of ater than dyacorum, with most of septum very narrow, broadening only at base. However, in photos, septum did appear broader than in visual inspection. Intermediate noseleaf broad and swollen-looking, as depicted for ater but unlike depiction of dyacorum.
Treated here as ater until further evidence emerges.
Observations:
Only identified from trapped individuals.
Echolocation:
Fc = 153 to 160 kHz, very low amplitude.
Very
difficult to detect, even from 50 cm.
Calls of very short duration, less than 5 ms.
Because of the small sample size, and expected variation, it should be
assumed
there will be overlap with H.
sabanus.
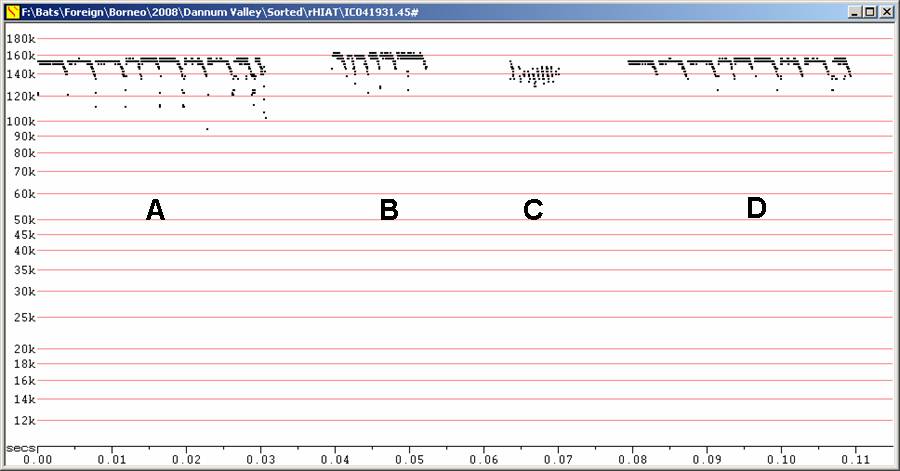
H155 at F8, compressed. Note high horizontal resolution.
A, B and D Recordings
from
hand showing variation in frequency
C
Recording from hand showing only partial detection of downsweeps
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from trapped individuals.
Echolocation:
Only recorded in hand. Very difficult to detect, even from 20 cm. Calls of very short duration (usually less than 5 ms). Recorded at frequencies between 147 and 153 kHz. Probably some overlap with H. ater.
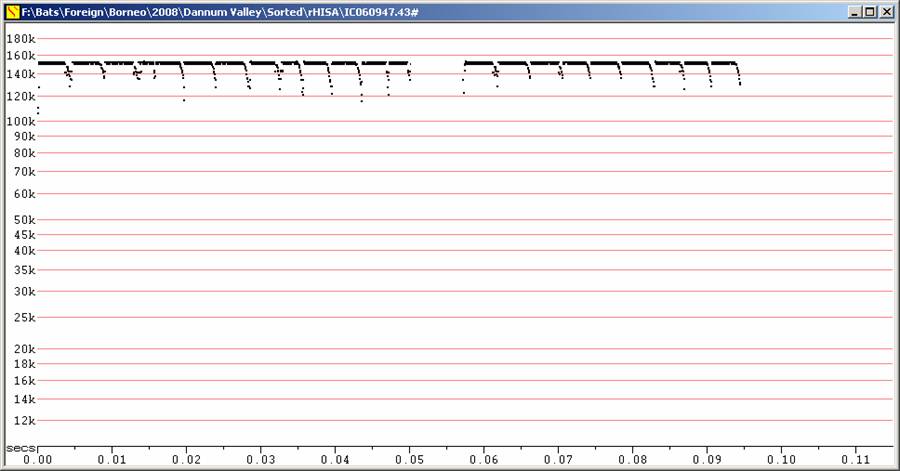
H150 at F8, compressed. Note high horizontal resolution.
ack to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Identified from captured individuals. Bats seen in the field giving calls like those depicted below were assumed to be H. cervinus.
Observations:
Commonly seen flying around dwellings at both Danum Valley and the Kinabatangan River. Often flew through brightly lit areas where insects collected under fluorescent lights. This was the bat most commonly seen hunting under verandahs and in close proximity to humans. Flight seemed fast and direct, turning abruptly and often very close to structures and people.
Echolocation:
Numerous field recordings at Fc = 111 to
118
kHz. Duration ca 7 ms. Recorded on release at 112 to 118 kHz, duration ca
4 ms.
Calls of low intensity, difficult to record from more than 2m.
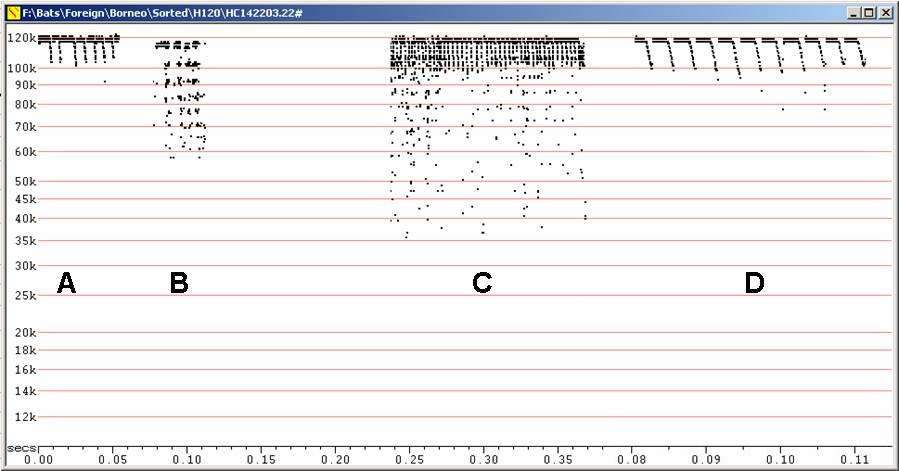
H115 at F6, compressed. D at F8, compressed (note change of scale).
A - In free flight in field. B - In field. C - In captivity.
D - Part of C expanded to F8.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from trapped individuals.
Echolocation:
Fc = 85 to 90 kHz, duration 30 - 80 ms
Frequency slightly higher than Rhinolophus borneensis, but expect some overlap.
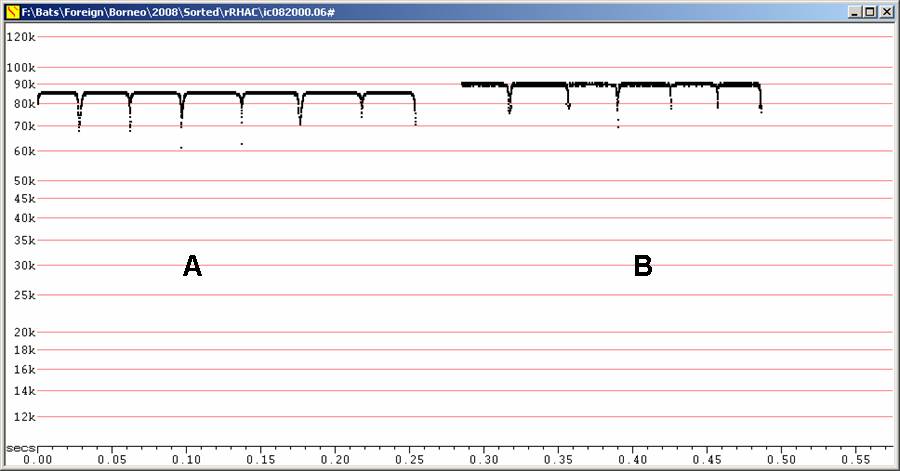
F6, Divratio 8, compressed
A - R. acuminatus recorded in
hand. B
- recorded in flight after release from hand.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from trapped individuals.
Echolocation:
Fc = 80 to 85 kHz, duration 20 - 60 ms
Frequency slightly lower than Rhinolophus acuminatus, but expect some overlap.
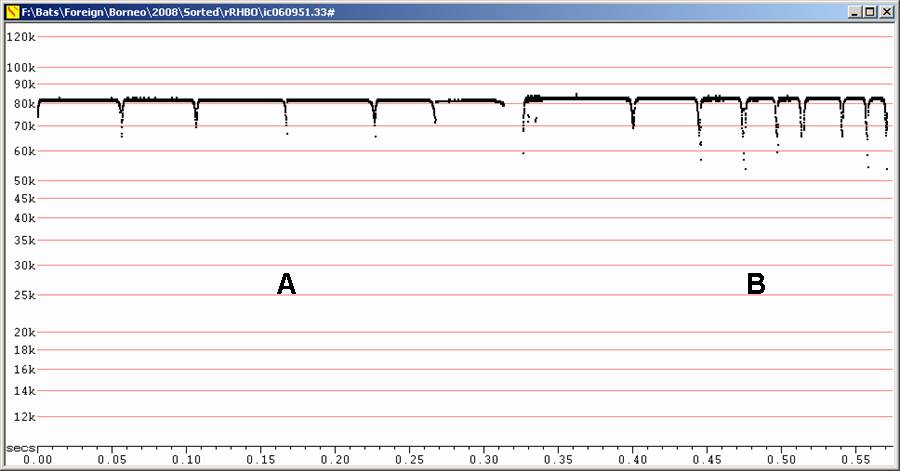
F6, Divratio 8, compressed
A - R. borneensis recorded in
hand. B
- recorded in flight after release from hand.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from one trapped individual.
Echolocation:
One individual captured and released. In
bag,
Fc = 67 kHz. In flight, Fc = 65 kHz, duration < 20 ms. Not recorded in
field.
Note similar frequency to Rhinolophus creaghi, but duration shorter and corners of calls more
sharply angled. Upsweeps difficult to see, but based on only one
individual.
H65, compressed. A and B at F6, C at F7 (note change of scale).
A - Calling from ceiling of walkway in building. B - After release. C -
Expanded
to F7.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from trapped individuals.
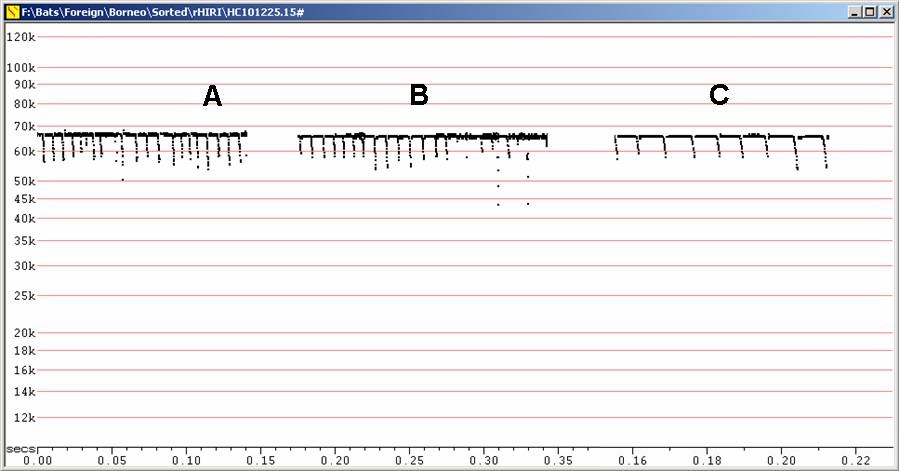
Echolocation:
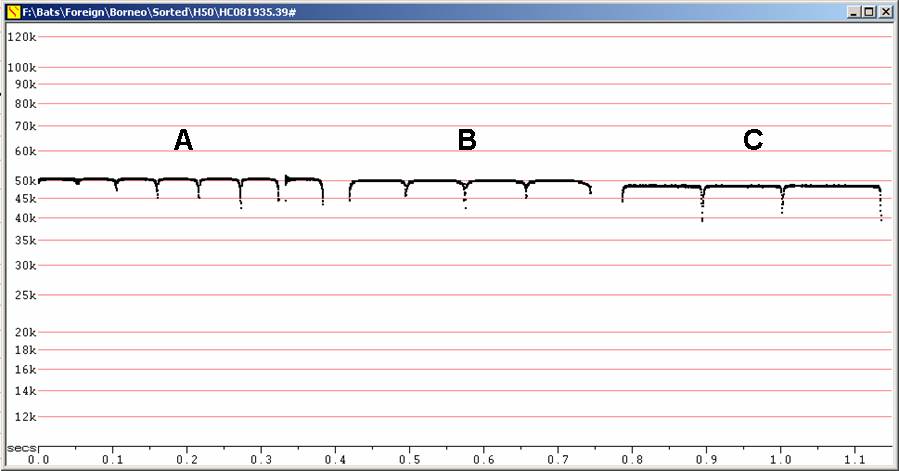
Fc = 63 to 65 kHz, duration 25 - 60 ms. Note similar frequency to H. ridleyi, but calls longer, more rounded corners and upsweeps obvious.
F6, Divratio 8, compressed
A - Recorded in hand. B - In flight after release from hand.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Identified from captured individuals. Bats seen in the field giving calls like those depicted below were assumed to be H. diadema.
Observations:
Several bats seen around the field centre at Danum Valley were considered to be this species on the basis of call and large size. Calls like those below were often recorded passively close to buildings, apparently from bats hanging in nearby trees.
Echolocation:
In bag, Fc = 64 kHz, in flight, Fc = 61 kHz, duration <20ms. Note frequency close to Rhinolophus sedulus, but shorter duration, terminal downsweep much more prominent than initial upsweep.
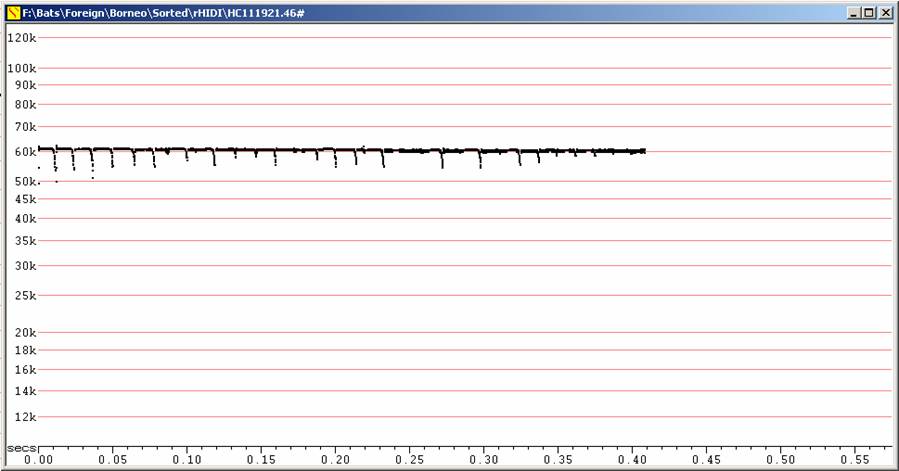
F6, compressed. In flight after release.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from trapped individuals. Bats attributed to this species (by call) were sometimes found hanging from vegetation in dark parts of heavily forested gullies.
Echolocation:
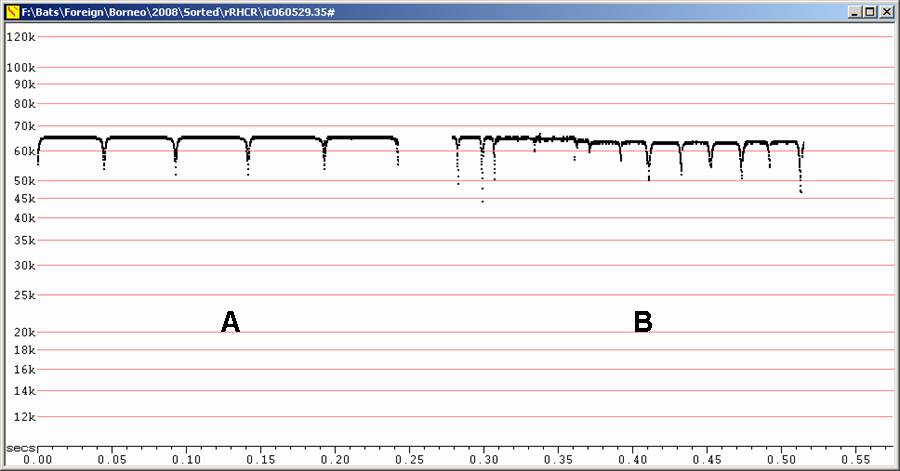
Stationary, Fc = 60 kHz, duration ca 50 ms. Not recorded in free flight. Frequency similar to Hipposideros diadema, but longer duration, initial upsweep like terminal downsweep.
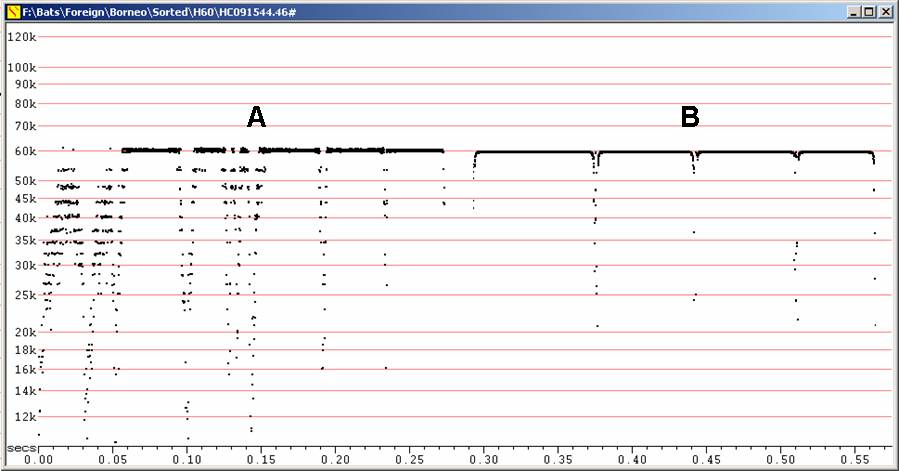
H60 at F6, compressed
A - Recorded poorly, showing breakup of
call.
B - Recorded well.
Both recordings from bats stationary, hanging from vegetation in gully
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Common around Danum Valley, with several captured in harp traps. Frequently recorded calling while hanging in trees close to roads or tracks. In that situation, usually seemed oblivious to nearby humans searching for them with lights. When found, they were usually quite exposed, hanging from branches over open spaces. It is unclear if these bats were calling in response to nearby humans, or were actively hunting while stationary.
Echolocation:
Fc = 48 to 50 kHz, duration > 30 ms.
F6, compressed.
A - In free flight. B - In hand. C - Same bat as B, recorded shortly after
release.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
A single individual was recorded while stationary in a tree, but not seen. Some digital cameras were found to produce 40 kHz signals from their autofocus mechanisms, but the calls depicted below look like real Rhinolophus, and one sequence shows drop in frequency consistent with a bat taking flight.
Echolocation:
Fc about 39 kHz, durations up to over 60 ms. The lower calls have Fc ca 38 kHz.
H38 at F6, compressed. B at F1, truetime (real time intervals for B are 40
times
those shown, so the sequence at B has a duration of nearly 7 seconds).
A - Three well-defined calls (F6, compressed). B - Poorly recorded calls
(F1,
Truetime) showing drop in frequency at end, consistent with a bat flying
from a
perch.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
One captured juvenile had FA = 42.4mm, Wt = 4.5g. Size suggests Emballonura monticola (MOB).
Observations:
Widespread in Danum Valley. Small/medium, pointy-winged, overall dark in colour. Tends to hunt high, usually close to or below treetops, usually fairly close to trees. Flight distinctive, jerky, regularly seeming to fold the wings in close to the body, so flight appears to occur in spurts. Commonly encountered along roadways, but rarely close to the buildings at the Field Centre.
Two species known from Borneo, E. alecto and E. monticola. Because of the wide range of frequencies recorded, it is possible that two species are present in the recordings from Danum Valley, but against this, most calls seemed to be about 45 kHz, in the middle of the range, rather than split between two frequency peaks.
Echolocation:
Echolocation very distinctive. Calls at between 41 and 50 kHz, on H2 (second harmonic). Often drops to H1 (first harmonic) at entry to or during attack and feeding buzz. May drop to H1 for just the droopy downsweep of each call in attack phase. Attack involves shortening call duration, eventually dropping the cap off the call so it becomes just a droopy downsweep.
Duration about 5 ms, TBC (Time Between Calls) ca 120 ms. TBC in buzz 5.6 ms.
N45 at F6, compressed
A
Sequence
with feeding buzz, followed by calls of different shape
B
Calls
showing abrupt drop in frequency at end
C
Higher
frequency example
D
Short
sequence with two bats at different frequencies
E
Sequence showing drop to H1 during attack/feeding buzz
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
In the genus Kerivoula, the following species were recognised:
K. papillosa KEPA
K. pellucida KEPE
K. hardwickii KEHA
Another species remains unidentified. It appears to be either K. intermedia or K. minuta, or perhaps a composite of both. We have no information which allow us to feel confident of distinguishing these species. In the rest of this text, this species will be referred to as K. sp, or by the abbreviation KEsp.
These four (or perhaps five) species are all very difficult to record, because their calls are so faint.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
A small Kerivoula with FA 30 to 32 mm. Dorsal colour a soft, grey-brown, without orange tones. Ventral fur with bases distinctly darker than tips, but without striking contrast. Ears a little longer than width of snout compared to KEsp. The ears look a little longer than the gap between their bases, and the tragus reaches to just over half the length of the ear.
Observations:
Only identified from trapped individuals.
Echolocation:
Extremely high frequency calls, with detection as high as 250 kHz. Only detected over very short range.
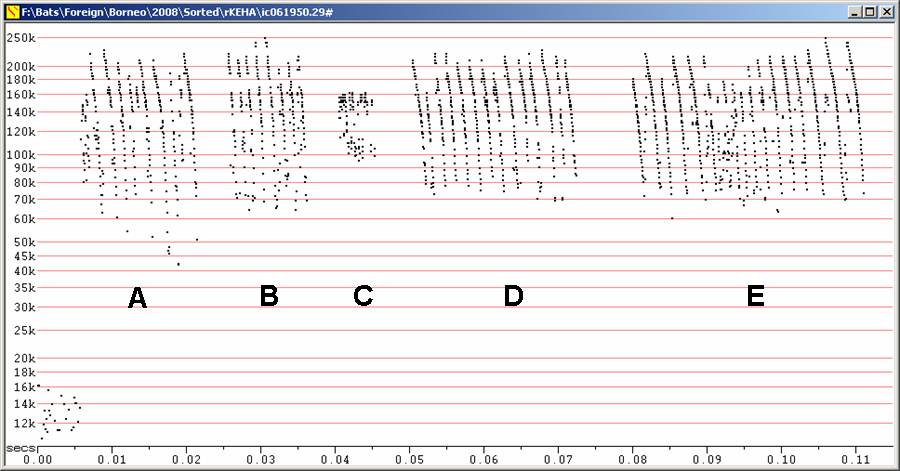
Kerivoula hardwickii at F8, compressed. Note wide range on frequency
axis.
A,B,C Calls recorded from
different bats released after capture
D,E Two
sequences
from one bat calling from hand just before taking flight.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from trapped individuals.
Echolocation:
Calls steep and of short duration, typical of the genus. Because of the wide range of call types given by this species (and the other Kerivoula recorded), it isn't certain how these species could be distinguished acoustically, or even if they can be distinguished from bats in the genera Phoniscus and Murina.
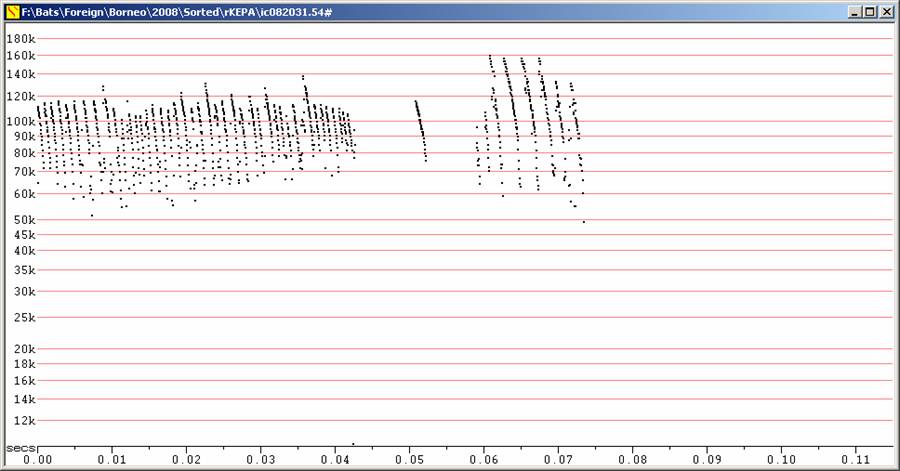
S100 at F8, compressed.
All recordings from bats released after capture, but not necessarily from
bats
in flight.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
A few individuals identified as of the genus Kerivoula were captured and released, but these were not identified to species. They were thought to be either K. intermedia or K. minuta, or possibly both.
Echolocation:
Like other Kerivoula, these gave
low
amplitude calls upon release which were quite difficult to record.
Some recordings were made of free-flying bats which could have been the
same
species, but these were brief, poorly-recorded
sequences.
The illustration below shows a lot of variation in call types, and of particular interest, that the same individual bat produced steep calls which reached to lower frequencies than the less steep calls. Note that these calls (A and B below) are consistent with Kingston's characterisation of K. intermedia or K. minuta calls (see Kingston et al, 1999. Echolocation Signal Design in Kerivoulinae and Murininae from Malaysia, J. Zool., Lond. 249: 359-374 ), though the differences are unlikely to lead to reliable identification because the variation is so great in this small sample. Note that even though the sequences in C and D seem lower in frequency than those in A and B, there were cases where the same individual bat gave calls resembling each type.
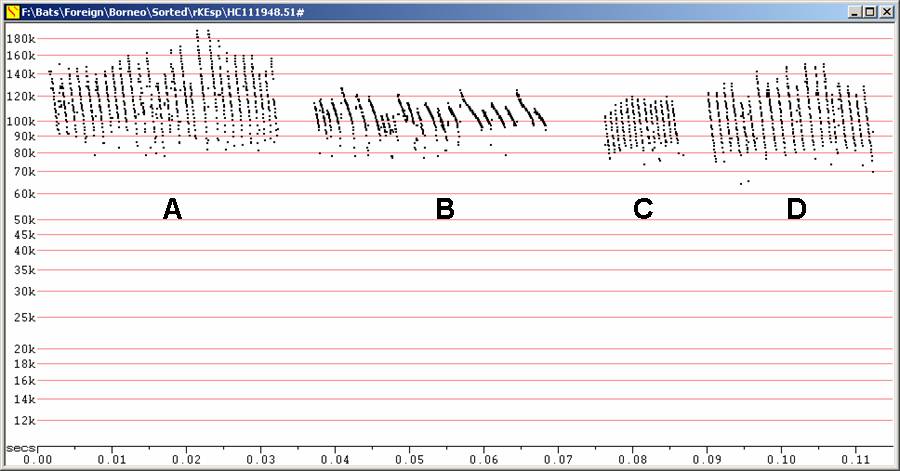
F8, compressed.
Note high magnification along time axis and extended frequency range
compared to
most other illustrations.
A, B - Two sequences from the same bat, released from capture
C, D - Two sequences from different bats released from capture
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
These bats were readily identified as Miniopterus by visual features and call type. They were quite evenly coloured without obvious pattern, and the wings looked narrow and the tail long. Flight fast and hard to follow. Looked very much like M. australis, though slightly lower in frequency than typical australis from eastern Australia. However, australis from Cape York are lower in frequency than those from elsewhere in Australia, and acoustically closely resemble these bats from Borneo.
Observations:
Uncommonly recorded, but at several locations along the road into Danum Valley, though not seen around the field centre itself.
Echolocation:
Note potential overlap with J50, but J50 duration typically markedly shorter. Visual observations showed a very different flight style and overall appearance.
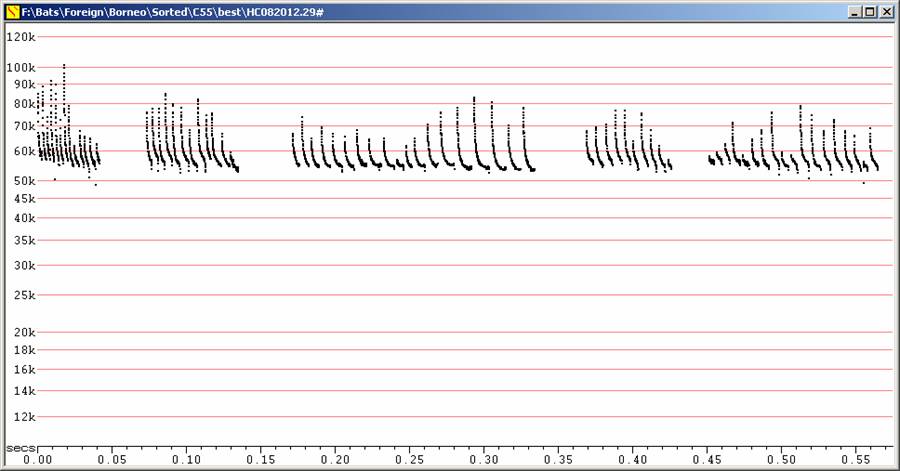
C55 at F6, compressed
Five different sequences recorded in free flight.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Only identified from trapped individuals.
Echolocation:
Two captured and recorded on release.
Low
amplitude, short duration calls, with all calls less than 1 ms in
duration, most
less than 0.5 ms.
Calls of this species appear to vary a great deal in minimum frequency,
from 50
to 90 kHz, but all calls recorded showed only a short sweep.
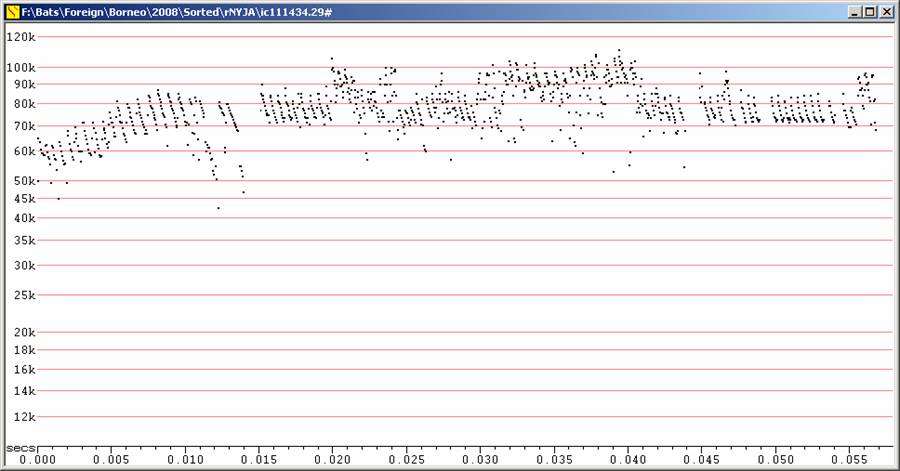
F9, Divratio 8. Note extreme magnification in time axis.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Water-skimming Myotis were seen occasionally at Danum Valley and commonly on the Kinabatangan River. At Danum, they were seen from above as they worked along a small creek inside the forest. At that site, the back colour looked strikingly pale compared to the wing membranes. This was not noted at the River, but there they were viewed from close to water level. The bats at the River showed a very distinctive flattening of the call at between 50 and 55 kHz. This was mostly absent from sequences at Danum Valley, but its presence in one sequence, suggests the same species may have been involved. It would not be surprising if the individuals at Danum Valley showed this feature less, as they were hunting in much higher clutter.
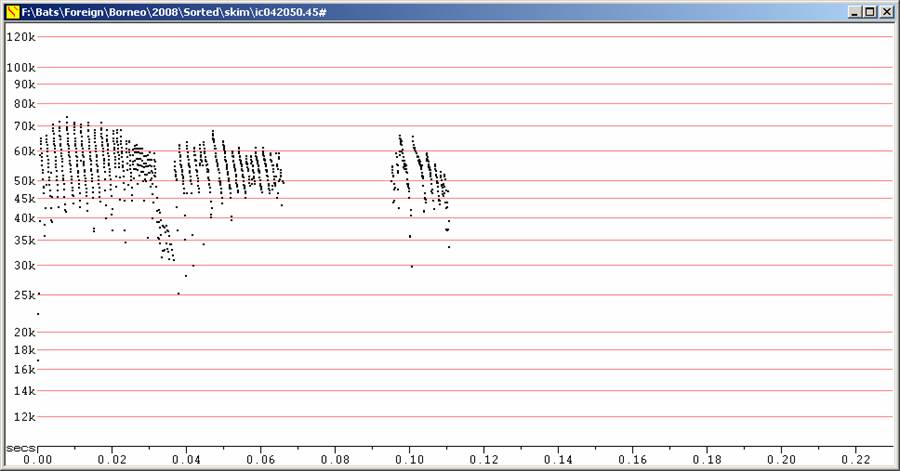
F7 at Divratio 8. S50 and B55 (or D55) at right.
Water-skimming Myotis at Danum Valley, including a feeding buzz. At right,
showing flattening of call at 55 kHz.
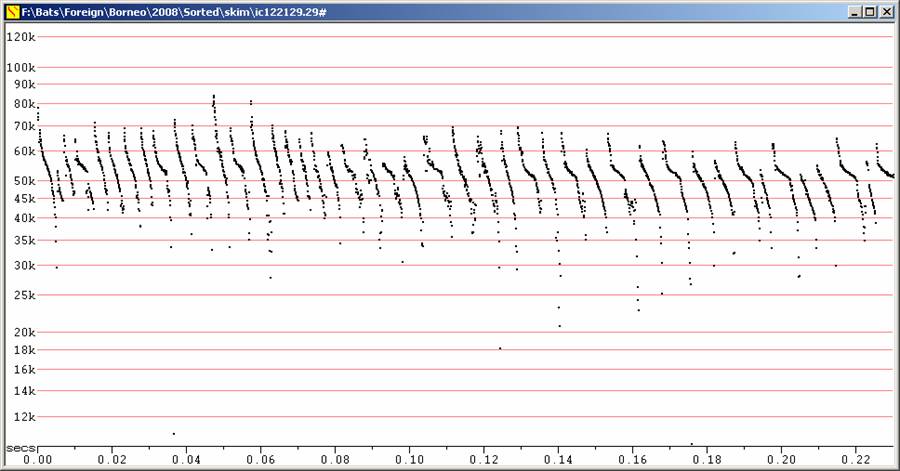
F7 at Divratio 8. Kinabatangan River. A mixture of call types from the one
individual, with Fc often about 50 kHz.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
One bat identified as this species was captured and recorded on release (see A below).
A few sequences of calls which might have been the same species were recorded from unseen, free-flying bats (for example, see B below).
It is quite feasible that the sequence at B could have also come from Myotis horsfieldii, with the obvious differences between the two sequences being quite consistent with the differences often seen between hand-released and free-flying Myotis. However, other species of Myotis could occur in the area, and Myotis are often difficult to distinguish acoustically, so there is no way to be sure at this stage.
Echolocation:
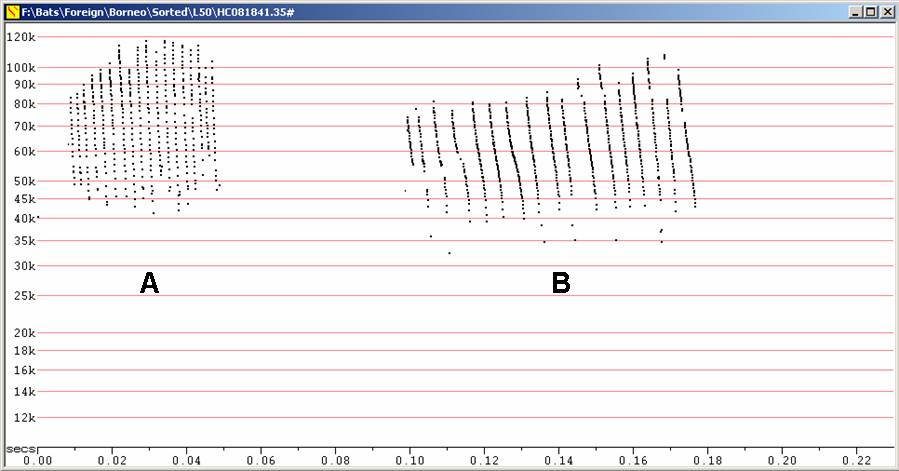
S50 at F7, compressed
A - Myotis horsfieldii
released from
hand
B - S50 calls recorded from unseen, free-flying bat.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
These bats were frequently captured and looked very typical of Myotis. They seemed to easily key out as M. muricola, based on MOB, although the teeth showed a lot of variability and were not all consistent with the published descriptions. Forearms were between 33 and 36 mm. In the hand, the captured bats showed two different colour variations, with most being strongly brownish in colour with an orange tinge to the abdominal fur. Some, however, lacked the brown tones and orange tints, and looked very grey all over. At least one of these was a juvenile, and the general appearance of these grey bats was consistent with juvenal Myotis. There was some variation noted in ear shape and head shape, but all of this seemed within the range of normal variation for a Myotis.
In 2008, Glenn Hoye considered that some of the bats captured were really M. ater. This raised doubt about the identity of these Myotis, and opened the possibility that two species were involved. There did not appear to be any consistent difference between the calls of brown versus grey bats. If these are different species, they would seem to be acoustically very similar.
Observations:
Many recordings from Danum Valley. Small, very fluttery, slow-looking flight - seemed to drift around rather than fly directly. One of the most frequently captured and recorded species at Danum Valley.
Echolocation:
Calls in the field typically J-shaped,
but
highly variable. Fc typically 48 to 53 kHz, but may be as low as 46 kHz.
Typical
duration about 5 ms, sometimes nearer 7 ms.
These calls are consistent with those of the bats captured and then
recorded
after release.
Acoustically, these bats are very unlike typical Myotis in Australia, North America or Europe, more like Central American Myotis. Their calls bear no resemblance to those of Myotis mystacinus, with which M. muricola was once lumped as the same species.
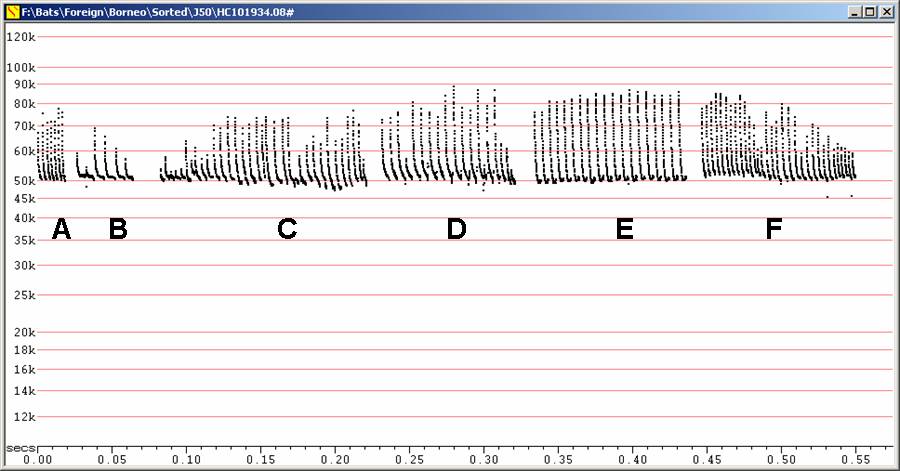
J50 at F6 compressed
A, B, C, D - Recorded in free flight
E, F - Myotis muricola / M. ater recorded after release from capture
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Bat not seen, and very few recordings.
Echolocation:
Short duration calls to about 6 ms and with only a short frequency sweep. These calls resemble calls from European species of Pipistrellus.
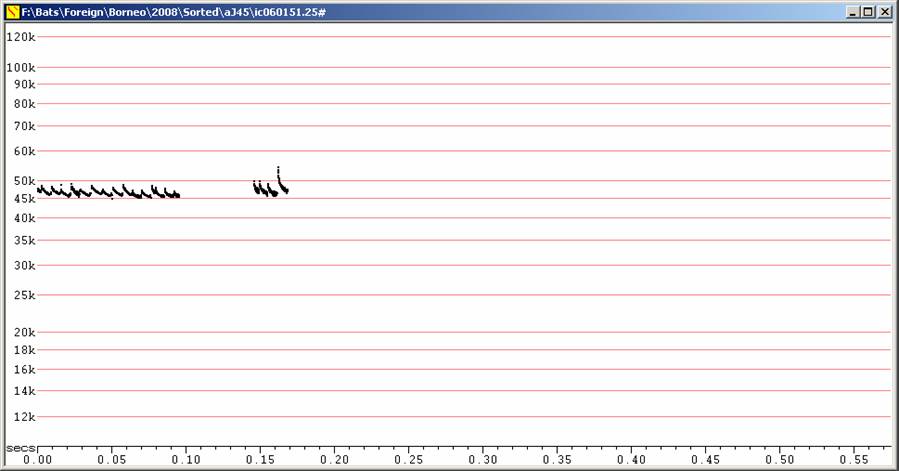
F6, Divratio 8, compressed.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Visual observations and acoustic characteristics were consistent with Miniopterus, and the feeding buzzes frequently observed are diagnostic of the Miniopteridae.
Observations:
The bat most commonly recorded in Danum Valley. On driving transects, did not appear in any numbers till later at night than most other species, but pretty well ubiquitous once it appeared. Often more than one present at once. Commonly observed around the buildings at the field centre.
This species was often seen hunting well below the canopy, with direct, fast flight. Looked like a Miniopterus, with long, pointed, narrow wings and long, pointed tail. Little colour pattern, but head often appeared darker than rest of body. Looked very like M. oceanensis in Australia, and calls similar, but lower in frequency.
Echolocation:
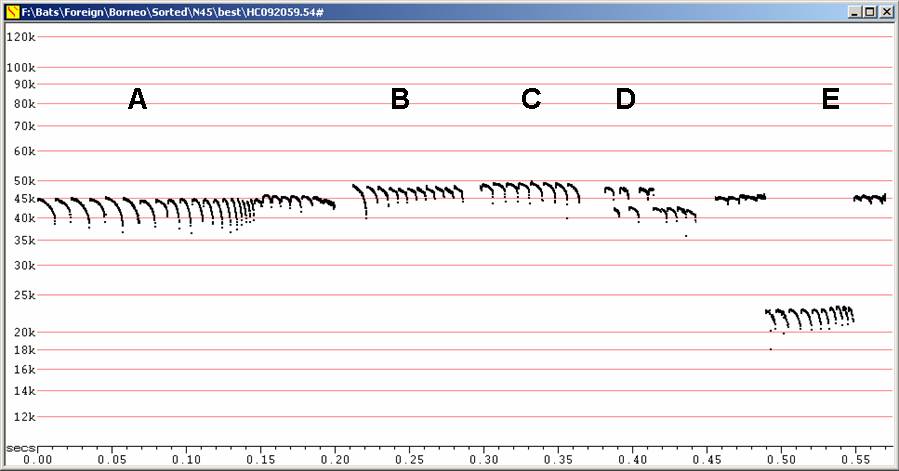
Moderate slope, curvilinear or bilinear calls dropping to 38 to 41 kHz, sometimes down to 37 or even approaching 35 kHz. Search phase calls may be J-shaped. Duration typically about 6 to 10 ms. Total sweep (difference between Fmax and Fmin) maybe as low as 5 kHz.
May show marked alternation in extent of call detected, suggesting head-turning.
Feeding buzz is diagnostic of Miniopterus, showing an abrupt drop in maximum frequency and a lack of the Buzz 1 phase. See Corben, Chris 2010. Feeding Buzzes. The Australasian Bat Society Newsletter. Number 35, November 2010. Pp 40 - 44.
May show jump to H2 late in pulse, especially in feeding buzz. Because of harmonic jump, buzz pulses may deceptively appear to start at much higher frequency than is real, and this could mask its distinctive nature.
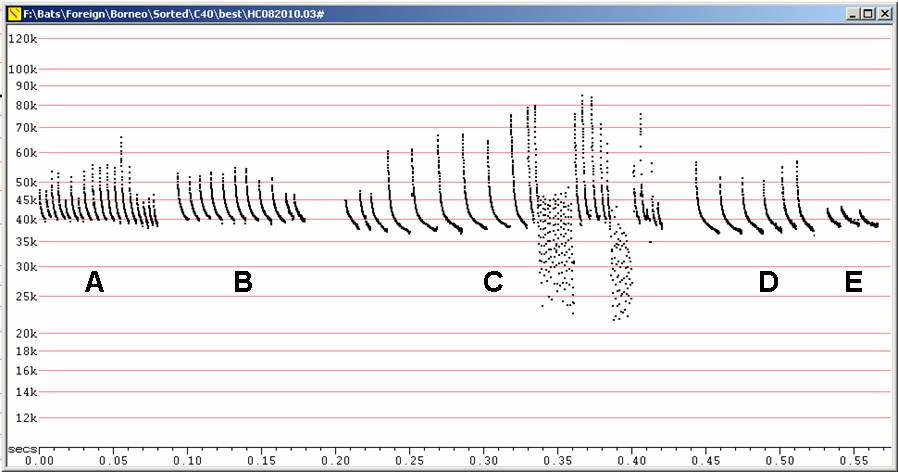
C40 at F6, compressed
A
Calls
in moderate clutter
B
Calls
in lower clutter
C
Two
feeding buzzes, showing sudden drop typical of Miniopterus.
D
Lower
clutter calls
E
A short series of calls in low clutter
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Uncommonly recorded, and not seen.
Echolocation:
Most calls curvilinear, but lower clutter calls distinctly J-shaped. Marked frequency alternation in some sequences.
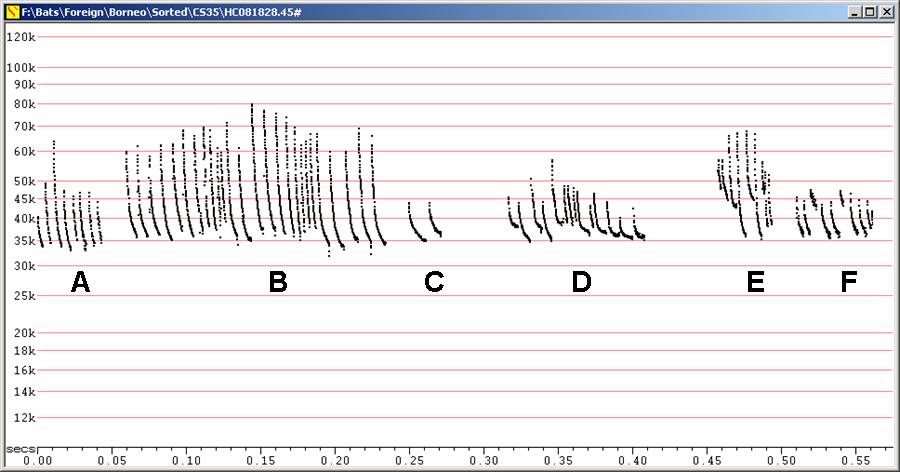
J35 at F6, compressed
A, C
Search phase
B
Search phase with attacks
D, E, F
Showing alternation
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Observations:
Not commonly encountered, and never seen.
Echolocation:
May produce F28 calls, which can easily be confused with F27, but frequently alternates by up to 8 kHz. Higher clutter calls are steep and higher in frequency, S35, B33 etc. These features suggest a Vespertilionide. Call frequencies consistent with Pipistrellus stenopterus (KLA).
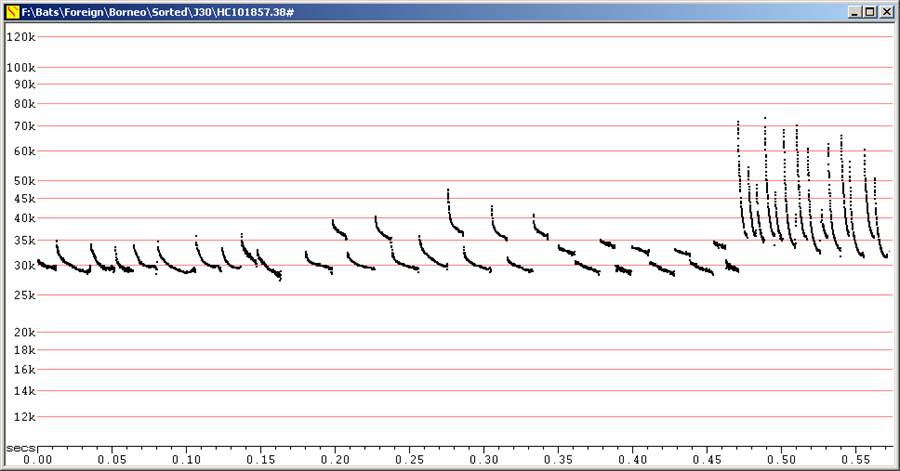
F28 at F6, Divratio 8, compressed
Four sequences showing variation in call shapes. Prominent frequency
alternation
in most sequences, including higher clutter sequence at right
.
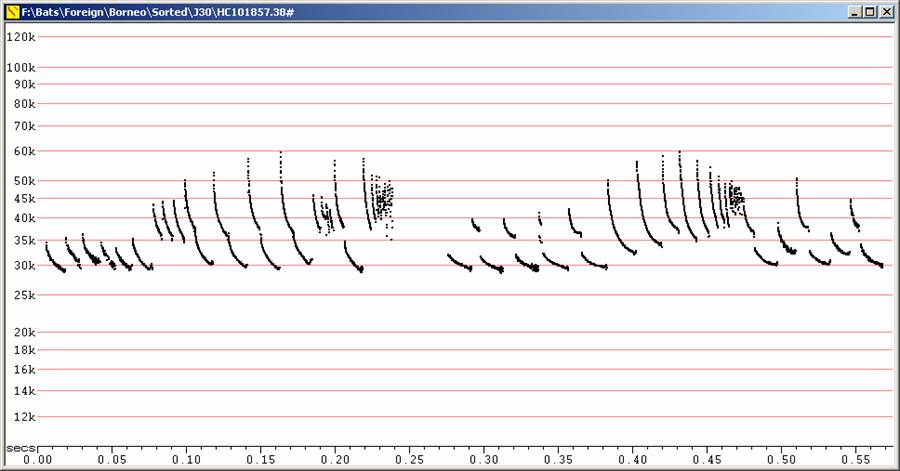
F28 at F6, compressed
Two different feeding buzz events. Note marked frequency rise into buzzes,
preceded by conspicuous frequency alternation.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
The calls recorded from this species show many features consistent with Molossides. Furthermore, visual observations showed a bat which is very similar in appearance to other known Molossides.
Of the Molossides known from Borneo, both Mops mops and Cheiromeles torquatus are elsewhere tentatively associated with very different call types to these F23 calls. Furthermore, Cheiromeles torquatus is easily eliminated by its much larger size.
In light of the above, the most likely identification for F23 is Tadarida plicata.
Observations:
Visually, looked very like Tadarida brasiliensis from North America. Quite small, pale below, looking quite white from some angles. Plain, with no obvious colour pattern. Wings narrow and pointed at tips. Flight fast, with rapid, continuous wingbeats, often recalling a Swift. Flight often erratic, with rapid changes in direction, often swooping. Always seen high above the ground, and often apparently out of visual range, as typically very difficult to see with a spotlight. Sometimes seen disappearing behind low clouds.
Numbers highly variable at DVFC, with some nights when this was the most frequently recorded bat, while on other nights there were very few. Often appeared in numbers, with several bats recorded at once. This lead to confusing sequences containing calls from several bats, often covering a wide range of call types and frequencies. Behaviour and acoustic variability seems quite typical of some other Molossides, such as Tadarida brasiliensis.
Echolocation:
Acoustically very variable, with many calls steep (eg S30 or S25) or thickened at lower end (eg B30 or B25). The frequency range shown by the steeper calls suggests the flatter calls could vary from about F20 to F28, widely overlapping with the F22 and F27 calls attributed here to Emballonurides.
Low clutter calls look quite similar to F27 and F22, but usually lower than F27 and much shorter in duration than calls of similar slope from F22. See notes on Emballonurides for acoustic characters separating Emballonurides from other families. Typical sequences from F23 include features such as presence of "grace notes" or feeding buzzes which easily distinguish F23 from Emballonurides.
Several examples were seen of attack phases in which the usual downsweeping calls were preceded by upsweeping calls. This is a very unusual pattern, though it may have social significance, rather than being a feature of attack phase itself. Curiously, I have only seen similar feeding buzzes in Chaerephon jobensis in Australia. Other attacks were seen without this feature. Attack phase seems to drop progressively into a feeding buzz in a manner quite unlike the feeding buzz of C20 and much higher in frequency than that of the D22.
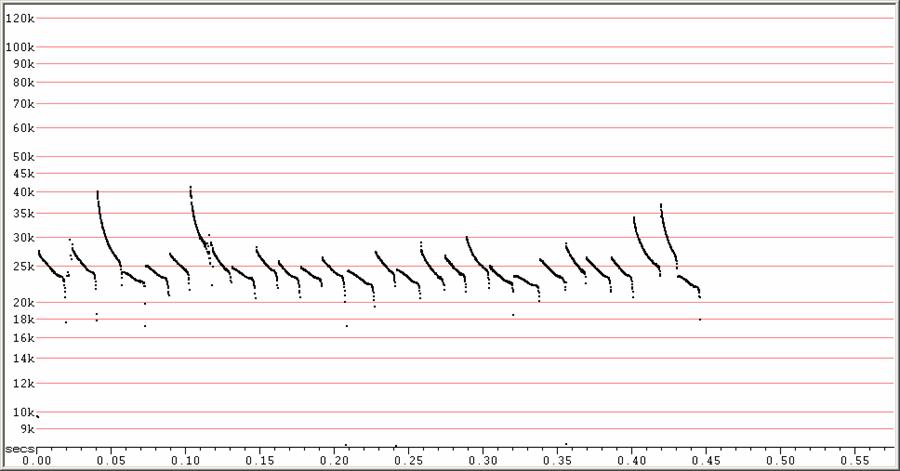
F23 at F6, compressed.
A typical search-phase sequence in low clutter. Note the higher, steeper "grace notes", a typical feature of Molossides and some Vespertilionides, but not of Emballonurides.
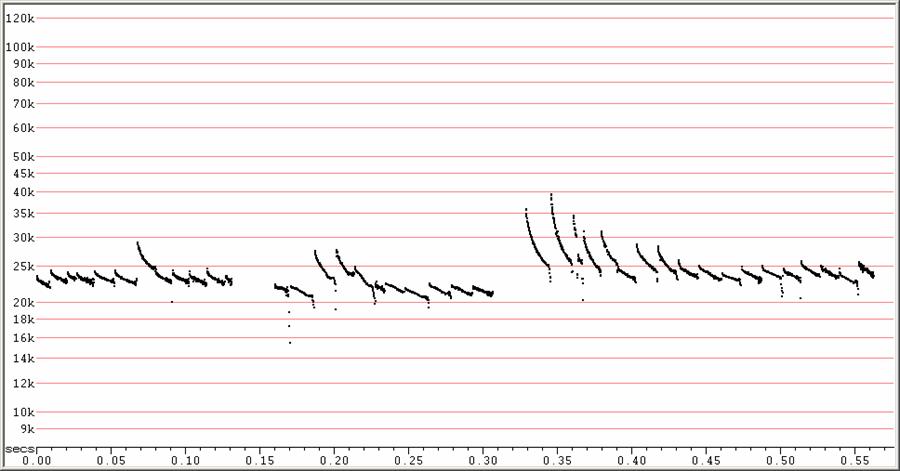
F23 at F6, compressed.
Search-phase sequences showing a variety of call shapes and frequencies. This variation is typical of many Molossides
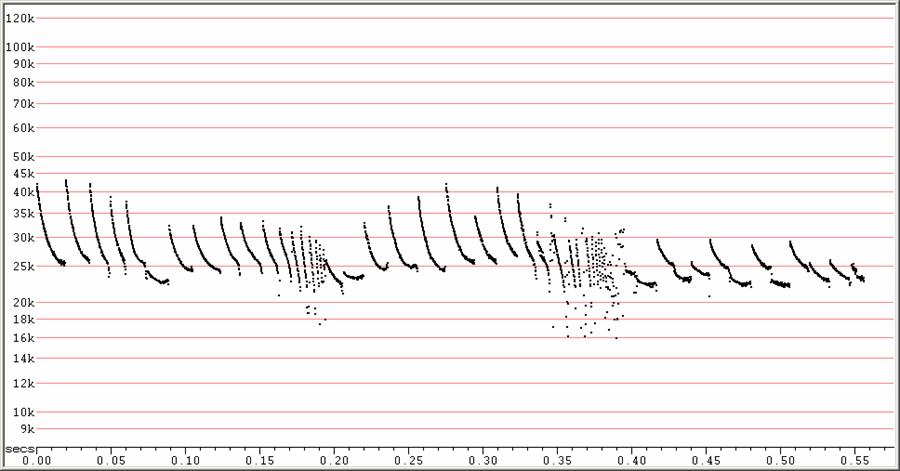
F23 at F6, compressed.
Rarer variant with upcurved, low clutter calls. Note pronounced frequency alternation at right. The higher clutter calls and feeding buzz seem typical for F23, and the curved low clutter calls are similar to variants often seen in other Molossides.
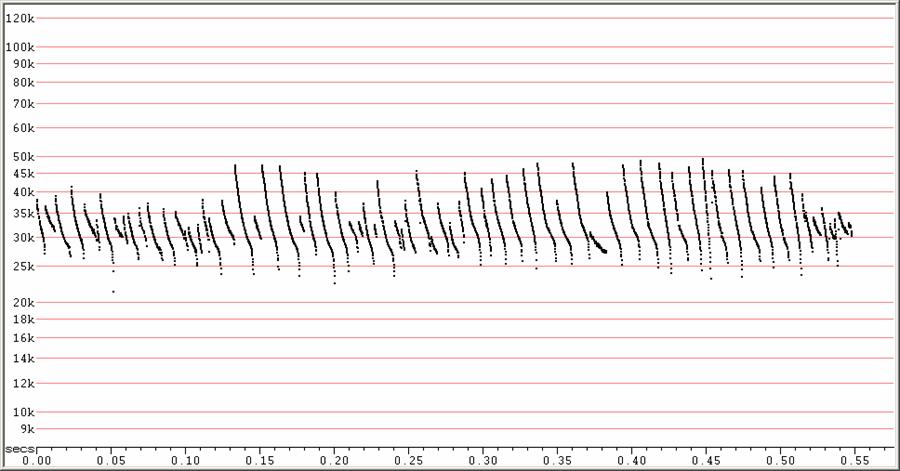
F23 at F6, compressed.
A variety of search-phase calls given by several bats flying in the same vicinity. The steepness of the calls suggests high clutter, but could also be a response to proximity to nearby conspecifics.
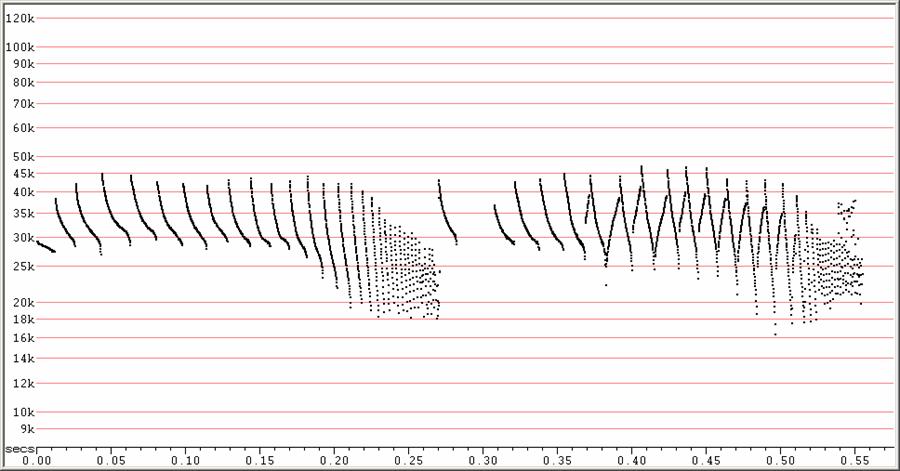
F23 at F6, compressed.
Two different feeding buzzes.
The buzz at right is an unusual variant in which the attack-phase pulses
are
preceded by pronounced upsweeps. This was seen several times in F23
sequences,
but most buzzes lacked this feature. This could merely mean that the
upsweeps
are too faint to detect from substantial distances. Note that these two
buzzes
would look very similar if the upsweeps went undetected in the second
case.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Two lower frequency call categories recognised here are attributed to Emballonurides, of the genera Taphozous and Saccolaimus, and certainly open space aerial insectivores, not members of the genus Emballonura.
Many calls of these species may be easily confused with calls of some Molossides or Vespertilionides. The most likely confusion in this area will come from the F23 calls of putative T. plicata flying high and straight in zero clutter. Emballonurides are usually only detected on the second harmonic, so any detection of part of a call on the first harmonic (at half the frequency) or the third harmonic (at 1.5 times the frequency) is clear evidence that the bat is an Emballonuride rather than a Molosside or Vespertilionide. However, detection of other harmonics is rare, so most sequences of these Emballonurides will be similar to sequences which could arise from other species.
Emballonuride calls don't differ much over a range of clutter, and their feeding buzzes are relatively inconspicuous, so any steep calls with long downsweeps will indicate other families. Most of the low slope calls given by other families will be associated with steeper calls, feeding buzzes or other features not given by Emballonurides, but the problem arises when bats are flying straight and high in zero clutter, when Molossides or some Vespertilionides can give long, consistent sequences of calls resembling those from the Emballonurides.
Bats of the genera Taphozous and Saccolaimus can be recognised in flight as Emballonurides by their wing and body shape.. Their wings are tapered to fine points like some Molossides, but are broad at the base, giving them a distinctive, triangular planform, unlike the overall narrower wings of most Molossides. The tail is also long, whereas tails on Molossides look short. Larger aerial Emballonurides show a slow, birdlike wingbeat, but smaller species look more normal in this regard, and Molossides show a parallel variation.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
The call features of this species clearly show it to be an Emballonuride. Known possibilities in Borneo are Taphozous melanopogon, T. longimanus and Saccolaimus saccolaimus. Other, lower frequency calls are attributed here to S. saccolaimus, so either of the Taphozous is likely to account for the F27 calls.
The F27 calls closely match the description in KLA for T. melanopogon. However, calls of several species of Taphozous in Australia appear not to differ much between species, so it is possible the calls described here as F27 could apply to either of the Taphozous, or perhaps both species have been recorded and assigned to the same call category.
Observations:
This bat was rarely observed visually, but many good views were obtained along the Kinabatangan River in 2009. It was occasionally recorded at Danum Valley, but was easily the most often recorded species in the city of Kota Kinabalu.
In colour and general form, this species looks like a small version of F22, with long, tapered, pointed wings, a long tail and overall white colouration, including the underwings and undertail. When flying away from the observer, the outer wingtips contrasted somewhat with the rest of the underwing, giving the impression of darker wingtips, but this effect disappeared once the bat was seen more directly from below. There was no sharp contrast between the underbody and the base of the underwings.
Compared to F22, these bats seemed even brighter white below, but with more contrast to the wingtips when seen at least partially from behind. The tail also looked proportionately longer. However, these differences could be just a consequence of the F27 bats providing better, closer views, as they typically flew much closer to ground level.
The flight action was extremely different from F22. The F22 bats showed a slow, birdlike wingbeat, and seemed to call once per wingbeat. The F27 bats showed a much faster, more typical bat wingbeat, with clearly more than one wingbeat between pulses when flying in the open. Note that the pulse repetition rate was similar between the two species under many conditions, despite the differences in wingbeat rate. The flight of the F27 bats looked faster and more erratic, often showing a certain jerkiness which was absent from the F22 bats.
Although it is extremely difficult to judge size of bats in any meaningful way, everything about the F27 bats made them look a lot smaller than the F22 bats, and there were many occasions when the bats were seen either in the same field of view, or very soon after one another.
Another way in which these species differed was that the F27 bats were usually seen in small groups of two to four bats, while the F22 bats were always seen alone, though recordings were made containing more than one F22 bat. The F27 bats seemed to be moving through the landscape in small flocks, so there would be periods when none were seen, then a small number would appear all at once and fly around together (but only very loosely connected - most often only one bat was detected at a time).
Along the Kinabatangan River, a night cruise showed that these bats were only recorded near small villages where there were lots of lights glowing. They were not encountered along the darker reaches of the river. At the lodge where we were based, the F27 bats were often first seen visually by the light radiating from bright lights placed near the edge of the river. These lights were as bright as typical, white streetlights.
Echolocation:
Commonly recorded as a sequence of low slope, long duration calls just above 25 kHz. A few sequences show traces of H1 at half frequency, or of H3, demonstrating that the calls usually seen are being detected on H2. This is consistent with this species being an Emballonuride. Frequencies are consistent with those reported for Taphozous melanopogon in KLA
In low clutter, durations could get up to 28 ms, though 20 to 25 ms is more common. Fc is about 26 kHz and Sc only 2 OPS. The total frequency sweep of such calls could be as low as 1 kHz. TBC for such sequences can exceed 1 s, though 300 ms or 600 ms is more frequent.
As clutter increases, the calls rise slightly in Fc, and in some cases, the initial downsweeps are replaced with upsweeps (F27-2_C). Higher clutter sequences may have Fc as high as 30 kHz, with TBC of 100 ms and duration often 10 to 12 ms (F27-2_D). In such cases, the initial and terminal downsweeps may be smoothly connected to generate a more curvilinear call.
In attack-phase, duration and TBC are reduced, but the frequency of the body is not changed much, though the initial downsweep can be lost and the terminal downsweep can be extended to as long as 7 kHz. These calls often appear as just a short body at the top of the call, followed by a conspicuous drooping tail. Attack-phase pulses can have durations down to 3 ms, and TBC can be down to about 7 ms.
Feeding buzzes are not often detected. Their pulses can have durations as short as 1 ms and TBC can get down to below 5 ms. These pulses can appear almost linear, though at high magnification, they still tend to show a generally drooped curvature. For this reason, the tops of the pulses are better defined than the bottoms, with Fmin appearing much more variable than Fmax.
There is no sign of frequency alternation or even of irregular jumps in frequency, except that the frequency rises when the bat moves into clutter, as indicated by a decrease in TBC. Therefore, there is a great deal of consistency in the calls given by a bat flying high and straight.
Compared to the F22 bats, the F27 bats were calling with much lower amplitudes. This was evident because they were much harder to detect, despite being closer to the observer, and also having calls to which the Anabat detector is more sensitive. Although the slightly higher frequency of the F27 calls would result in more atmospheric attenuation, this difference should be slight, and there was possibly even overlap in call frequencies.
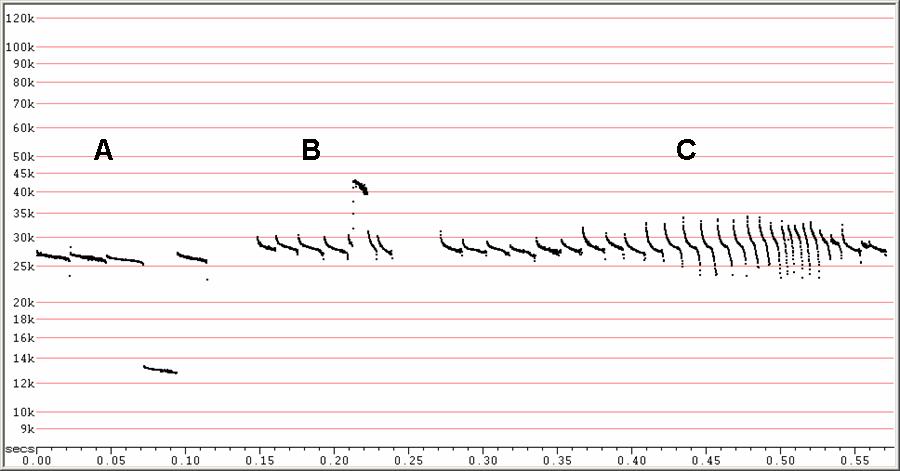
F27. F6, compressed.
A) Shows low clutter, search-phase calls with one pulse on H1 only.
B) One call jumping up to H3 partway through the pulse.
C) Shows typical search to attack-phase transition.
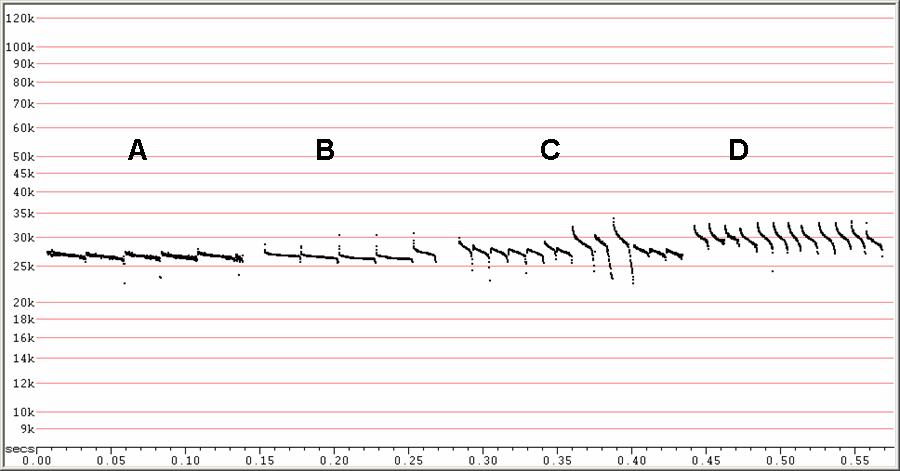
F27. F6, compressed.
Four types of search-phase sequences. Clutter increasing from left to
right.
A) Linear calls detected from bat in the open.
B) Linear calls with prominent initial and terminal downsweeps. These
features
might be present in the calls at A), but not detected.
C) Showing initial upsweeps instead of downsweeps.
D) More curvilinear form from bat in higher clutter.
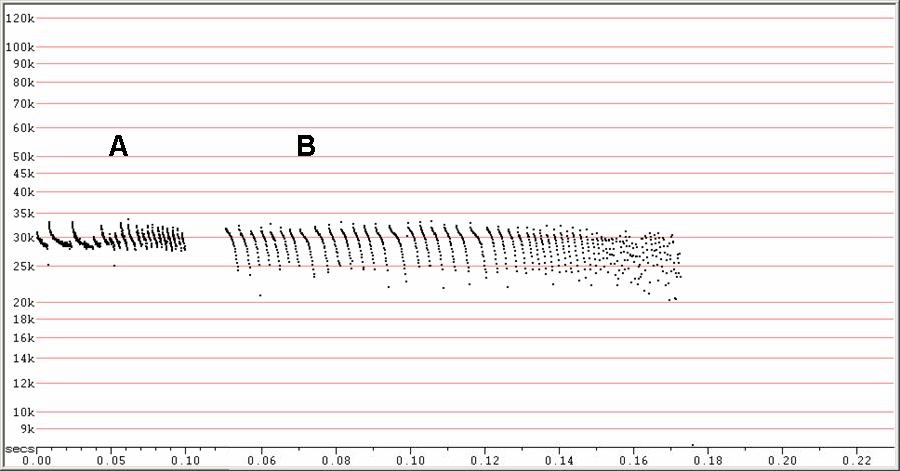
F27. A) F6, compressed. B) F7, compressed (note change in time scale).
A) Variant attack-phase, with less downsweep showing, probably due to
poorer
detection.
B) Expanded view of a prolonged attack and feeding buzz.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
Visual observations show a large bat with birdlike wingbeats and strikingly white underparts. The underwings also looked very pale or even white in many views. The bat was always seen flying high above the ground (or water), and sometimes passing behind low clouds. Overall appearance recalled S. flaviventris from Australia.
Echolocation calls were unmistakeably those of an Emballonuride, showing traces of a lower first harmonic in some recordings, and showing typical Emballonuride feeding buzzes. This species was therefore an Emballonuride with white underparts, which is apparently typical of S. saccolaimus but possible also on Taphozous melanopogon. Call frequencies are inconsistent with T. melanopogon as detailed in KLA, and also lower, which would fit with S. saccolaimus, which is markedly larger than T. melanopogon. Echolocation characteristics, including frequency, are also consistent with S. saccolaimus in Australia.
Observations:
Commonly encountered at DVFC, however, possibly few individuals involved. Hunted high overhead on dark nights, but seemed to be absent during periods when the moon was bright. At the Kinabatangan River, was commonly encountered hunting high over the river, but only in the vicinity of villages where there was extensive lighting. On a night cruise along the river, none were encountered in the dark parts of the river. They were frequently seen in the light emanating from the villages, and could then be watched with a spotlight. They did not appear to be shy of light, and when illuminated, continued without altering course or call type.
F22 bats were observed catching insects several times, most often with a short, rapid swoop downwards from typical flying height. Sometimes they would swoop upwards to attack an insect, and sometimes they would swoop down in a very long dive.
In colour and general form, this species looks like a large version of F27, with long, tapered, pointed wings, a long tail and overall white colouration, including the underwings and undertail. When flying away from the observer, the outer wingtips contrasted slightly with the rest of the underwing, giving the impression of darker wingtips, but this effect disappeared once the bat was seen more directly from below. There was no sharp contrast between the underbody and the base of the underwings.
Compared to F27, these bats seemed a little less white below, and with less contrast to the wingtips when seen from behind. The tail also looked proportionately shorter. However, these differences could perhaps result just from the fact that the F22 were always further away, allowing less detail to be seen.
The flight action was extremely different from F27. The F22 bats showed a slow, birdlike wingbeat, and seemed to typically call once per wingbeat. The F27 bats showed a much faster, more typical bat wingbeat, with clearly more than one wingbeat between pulses when flying in the open. The flight of the F22 bats looked more powerful and purposeful, without the more erratic and jerky aspects seen in the F27 bats.
Although it is extremely difficult to judge size of bats in any meaningful way, everything about the F22 bats made them look a lot larger than the F27 bats, and there were many occasions when the bats were seen either in the same field of view, or very soon after one another.
Another way in which these species differed was that the F22 bats were usually seen alone, though recordings sometimes contained more than one F22 bat. The F27 bats seemed to be moving through the landscape in small, loose flocks of two to four.
Echolocation:
Commonly recorded as a sequence of low slope, long duration calls just above 20 kHz. A few sequences, usually of higher clutter calls, show traces of a first harmonic (H1) at half frequency (about 12 kHz), demonstrating that the calls usually seen are being detected on H2. This is consistent with this species being an Emballonuride.
In low clutter, may produce almost linear calls of very low slope (eg 2 OPs), long duration (usually over 30 ms) and dropping in frequency by only 1 or 2 kHz, with Fc around 19 to 21 kHz. A few, poorly-recorded sequences suggest that Fc might even get as low as 17 kHz. Low clutter sequences may show consistent frequency, so they appear as a very regular sequence of similar calls, often spaced at about 350 ms intervals. More often, they show frequent changes in frequency, sometimes in the form of regular frequency alternation, and sometimes forming triplets of descending frequency. In such cases, the higher frequency pulses are associated with shorter TBC intervals, often about 200 ms.
In more cluttered circumstances, calls typically show a short initial downsweep and a longer terminal downsweep. The TBC is still 300 ms, but slope increases to about 5 OPS and the duration may drop to 25 ms, but is usually higher. Fc rises to about 22 or 23 kHz.
At higher clutter, the calls may become concave (as viewed from above) curvilinear, and the Fc can rise to 23 to 24 kHz. TBC may drop to less than 200 ms and the slope between 5 and 10 OPS. Duration is usually still about 30 ms, though the longest calls seen are of this type, with durations up to 40 ms. Maximum frequency at the start of the call could be above 28 kHz.
In an attack sequence, the calls get progressively shorter in duration, down to 5 ms, but with only a slight increase in Fc to about 25 kHz. The initial downsweep is progressively reduced but the terminal downsweep increases till the minimum frequency may be as much as 7 kHz below Fc. TBC drops to 20 ms. A following feeding buzz can show TBC down to 8 ms, with pulse duration down to 2 ms. The pulses in the buzz show reduced frequency range, and converge towards being linear, but still show a tendency to convexity - ie, the slope increases after being a minimum at the start of the call.
Both the attack-phase and feeding buzz are dominated by pulses which start with a low slope then droop downwards with increasing slope.
Compared to the calls of F27, F22 bat calls were lower in frequency for the same call types. They also showed a much greater variability in frequency, often taking the form of regular frequency alternation or irregular jumps in frequency without much change in call shape.
Compared to the F27 bats, the F22 bats were calling with much higher amplitudes. This was evident because they were much easier to detect, despite being much further from the observer, and also having calls to which the Anabat detector is less sensitive. Although the slightly lower frequency of the F22 calls would result in less atmospheric attenuation, this difference should be slight, and there was possibly even some overlap in call frequencies.
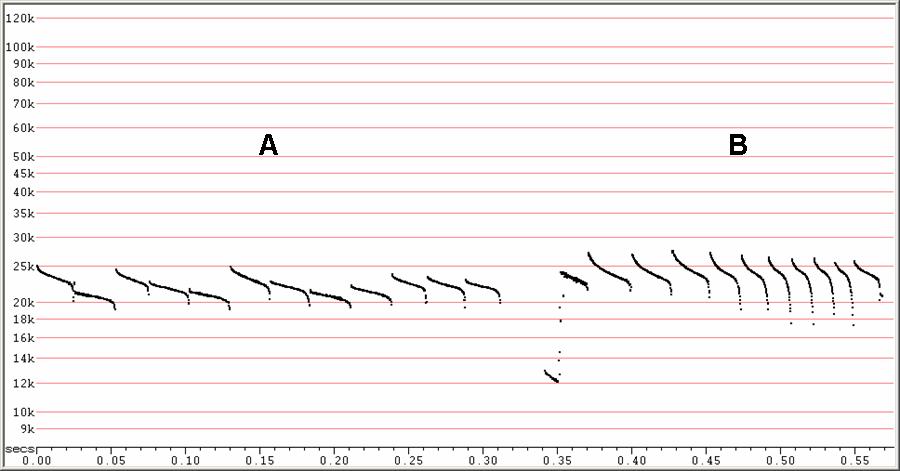
F22 at F6, compressed
A) Typical search-phase sequence, showing irregular changes in frequency.
B) An attack sequence with the first pulse showing H1.
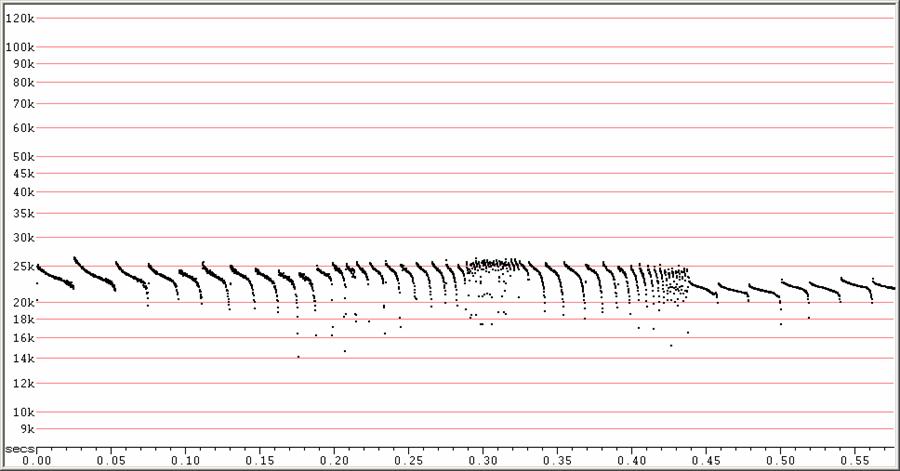
F22 at F6, compressed
Two attacks and feeding buzzes, followed by search-phase calls. All from
one
sequence.
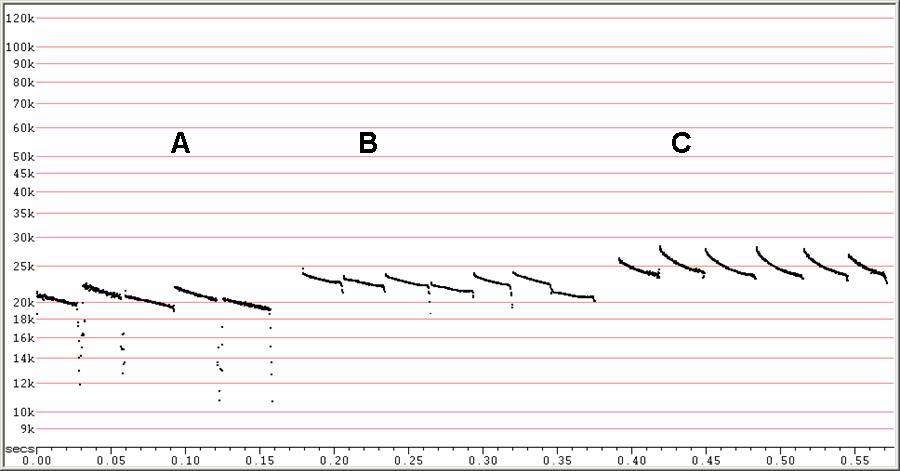
F22. F6, compressed
Three different types of search-phase sequence, clutter increasing from
left to
right.
A) Showing frequency alternation.
B) Showing some irregular variation in frequency
C) Showing consistent frequencies in the concave calls at right.
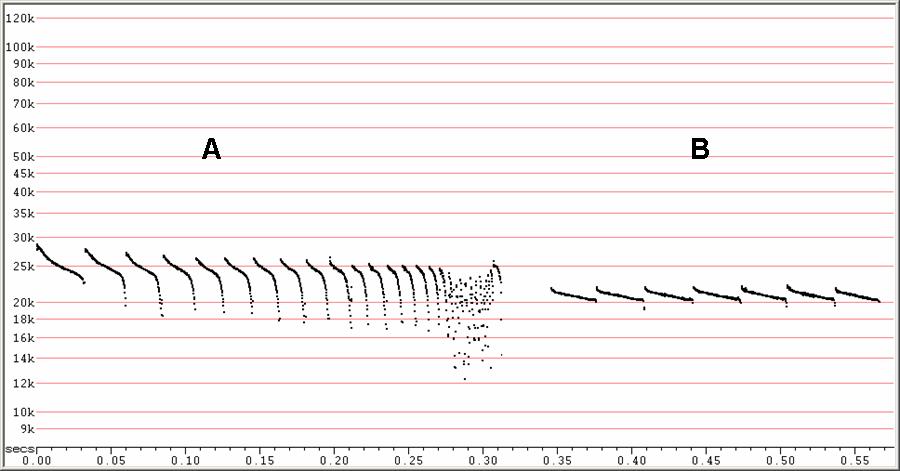
F6, compressed.
A) Attack and feeding buzz.
B) Search-phase sequence without frequency variation.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
The large size and unusual shape and calls suggest that this species is Cheiromeles torquatus. Other, similar-sized echolocating species known from Borneo are in the genus Taphozous or Saccolaimus, but the D22 bats looked and echolocated quite differently from Emballonurides. Furthermore, Cheiromeles is claimed to have an exceptionally high wing-loading, which is consistent with the way these bats appeared in the field.
The calls described here seem to differ from those depicted in KJAK. However, this may be more an artifact of the way the calls are displayed. A careful examination of Fig 2b of KJAK reveals that the Sc of the higher pulse is actually lower than that of the lower pulse - consistent with what is shown here. I feel the differences can be reconciled when the range of call types observed is considered. In any case, there is no credible alternative identification for the D22 bats.
Observations:
Commonly encountered, especially early in the evening. Visual observations showed a large, dark-coloured bat with slow, birdlike wingbeats, and unusually long, narrow wings, which combined with a very short tail, recalled Silver-rumped Swift Rhaphidura leucopygialis in overall shape. Size very large for an echolocating bat, recalling Eumops perotis of North America. The shape was very distinctive and unusual, not closely resembling that of other bats.
These bats were not seen flying very high above the ground. While it was obvious that F22 bats tended to fly high, the D22 were only seen lower down, well below the tops of the trees. A good way to see this species at Danum Valley was to stand on the suspension bridge over the river soon after sunset. As the light faded, these bats were seen flying along the river course.
Echolocation:
Calls complex and highly variable, but usually shows a long downsweep from as high as 35 down to as low as 15, but this sweep usually shows marked changes in slope. The initial downsweep is often not seen but starts from as high as 35, then curves down into a body which may start as high as 30, but more often 25 - 27. Fc may be 20 to 24, but usually more like 22. After the body, there is a long, steeper downsweep which may drop as low as 15. This final downsweep may show a marked droop, but is often quite linear.
There may be a marked, flat toe cutting off the final downsweep at anywhere from 20 to 22 kHz, and this may even happen before the final downsweep starts. Similar flat toes can be seen at the bottom of the call after the terminal droop.
These calls are usually very distinctive and long in duration, even over 30 ms. However, this species also often alternates these calls with higher frequency calls. The higher calls are usually curvilinear or J-shaped, but may be bilinear, and typically drop to 26 - 30 kHz. They often show irregular changes in slope, and may have brief flat portions and droops at the end. These higher calls are not always alternated, and may even be detected without the lower calls. They are sometimes given more than once between low calls. They appear to represent a more interrogative phase, and could even be thought of as part of the attack phase, as they sometimes are seen to lead into the feeding buzz - ie the calls change over from low to high types during the attack.
When these bats are flying higher above the ground, the calls may seem to drop down in frequency, so the shape is similar but the frequencies are reduced. This effect could be due to an artifact, where the higher parts of the call have not been detected. An example appears in the second figure below, which shows calls dropping all the way to below 15 kHz. These calls may not be complete, missing higher frequency portions which could even look like some of the calls in the middle sequence. In any case, the nett effect is that the calls seem to drop in frequency as the bat gets further out of clutter.
This acoustic behaviour is very unusual and doesn't seem at all typical of Molossides in general. The calls are much higher in frequency than of those of the similar-sized Eumops perotis of North America. The most unusual feature is the long, often droopy downsweep following the flattest part of the call. In some ways, this seems reminiscent of calls given by some other species when in high clutter. This opens the possibility that these observations are missing a core part of the repertoire of the species. What will it call like when flying very high above the ground in zero clutter? It could be that such calls are at much lower frequencies and lower slopes than those witnessed here. It is even possible they are not detectable from the ground.
While most sequences are very distinctive, some could cause confusion with C20 or other species. It would be quite possible to confuse the higher frequency pulses with other species if they were detected without the presence of the lower frequency pulses.
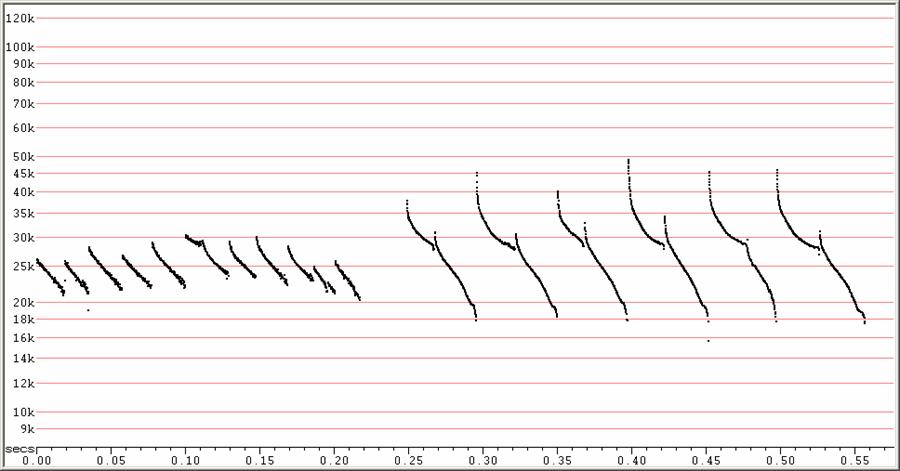
D22 at F6, compressed.
Two commonly encountered sequence types. At left, showing just one higher
frequency call amongst a series of lower frequency calls. It is possible
that
the appearance of this sequence is a result of the bat being too far away
to
detect the lower portions of the calls, in which case, the calls might
really
resemble those in the middle sequence below.
At right, a sequence showing obvious frequency alternation.
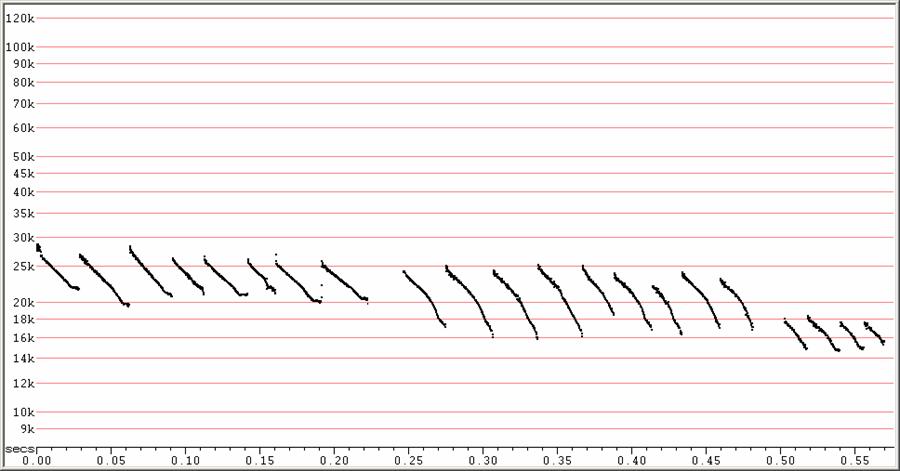
D22 at F6, compressed
Variations in search-phase calls. The sequence at right might actually
show just
the bottom portions of calls more like some of those in the middle
sequence.
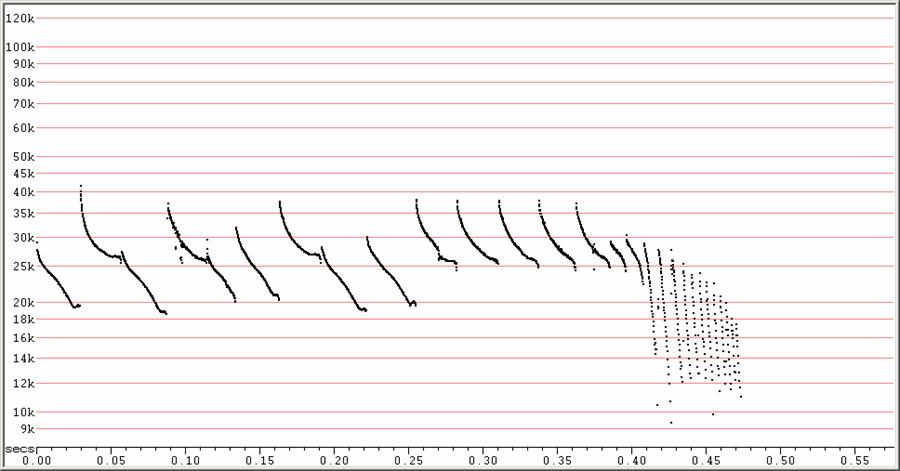
D22 at F6, compressed
Feeding buzz, with attack phase of high calls taking over from low calls
as
attack begins out of a frequency alternating search-phase sequence.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
General form and size eliminate most possibilities, leaving the larger Emballonurides, possibly a Molosside, Hesperoptenus tomesi and Scotophilus kuhlii, of the bat species known from the region. The long tail, broad wings and rounded wingtips all point to a Vespertilionide rather than one of the other families. The echolocation calls are wrong for Emballonurides, and putative acoustic identifications are suggested for all the Molossides expected. Call frequencies are way too low for Scotophilus kuhlii as detailed in KLA. The prominent frequency alternation is consistent with descriptions of calls of Hesperoptenus blanfordi in KLA, though the frequencies are much lower and the durations much longer, though this might be expected in a much larger bat. By a process of elimination, Hesperoptenus tomesi seems the most likely candidate.
Observations:
Several visual observations show a large (estimated forearm 50 - 60 mm, but this could be way off) bat with mostly dark brown fur, sometimes looking blacker on head. Noticeably larger than the C40 Miniopterus, which was often seen about the same time. Wings long, broad and rounded at the tips. Tail long, and looked squared at tip. One individual was strikingly white under tail, along flanks and in a broad patch under the base of the wing, but this feature was subdued or not apparent in other individuals, so may be aberrant.
These bats were seen flying across open spaces and often near crowns of tall trees, but always well above the ground. Nearly all visual observations were after the lights were turned out in the village, though the bats were recorded at other times. They seemed shy of light, seeming to move quickly away when illuminated by a spotlight.
Echolocation:
Produces a variety of call types, many showing profound alternation, with the higher pulses more J-shaped and very variable in frequency (ie 23 to 32). Lower pulses curvilinear and up to about 20 ms duration, with Fc from 19 to 24.
Some potential for acoustic confusion with D22, but vastly different bats in appearance and size, and similarities in echolocation only superficial. Pulses usually much shorter in duration than for D22, and not drooping downwards, but with slope still decreasing to 20 kHz, so concave from above rather than convex. Feeding buzz follows attack phase with rising frequency, unlike attack phase in F23, and higher in frequency than in D22.
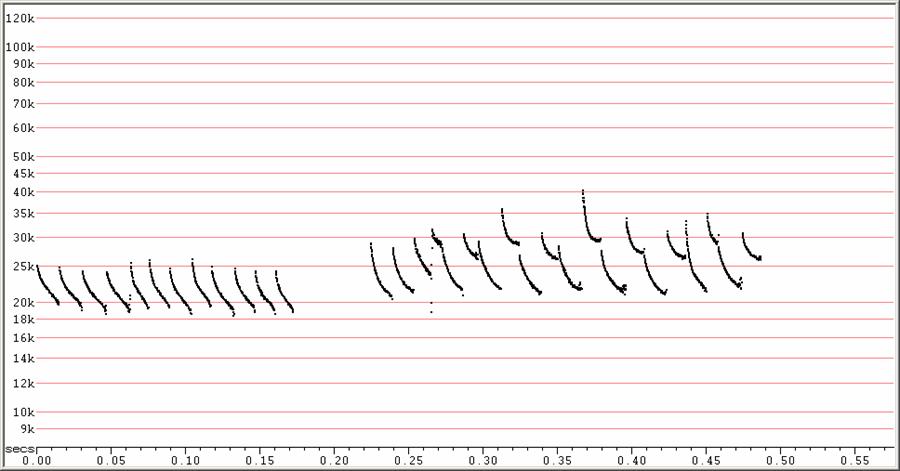
C20 at F6, compressed
At left, a sequence of calls without alternation and showing frequency
dropping
below 20 kHz. At right, a typical sequence showing conspicuous frequency
alternation.
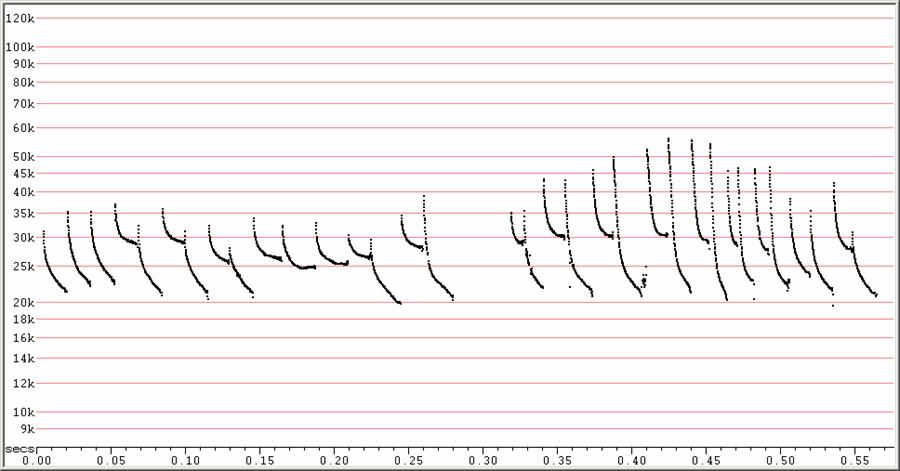
C20 at F6, compressed
Two sequences showing frequency alternation. At left, showing a sequence
in
which higher frequency pulses are given without lower frequency pulses. At
right, a sequence suggesting higher clutter, showing steeper, shorter
duration
pulses.
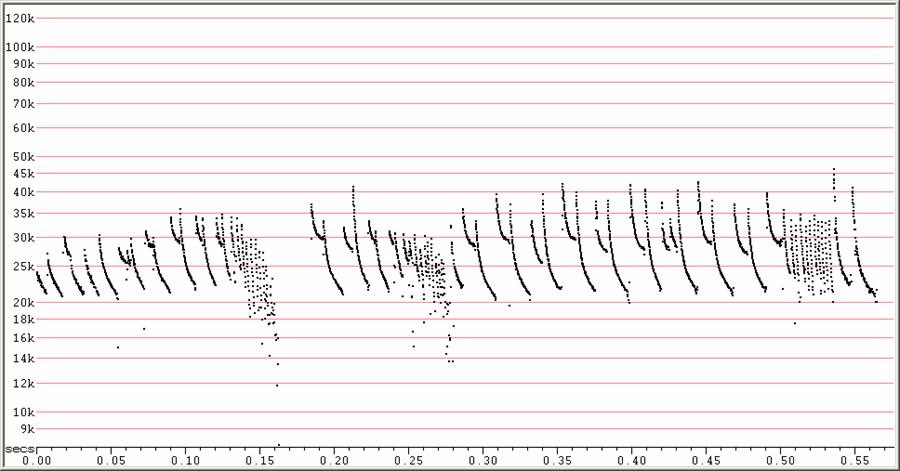
C20 at F6, compressed
Feeding buzzes from two individuals. A single sequence with two buzzes at
right.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
Basis:
The general acoustic behaviour of this species is consistent with a Molosside and not with an Emballonuride, because of the strong frequency alternation with the higher calls of very different shape, and because the attack-phase calls show a marked change in slope and frequency from the search-phase calls. The frequency is consistent with Mops mops as detailed in KLA, though showing smaller frequency sweeps, but that would be consistent with a bat flying more in the open than those producing the calls described in KLA. There are few species known from Borneo which could call in this frequency range, so the ID is based largely on a process of elimination.
Observations:
Not seen.
Echolocation:
In open, produces sequences of long duration (up to 20 ms), low slope (5 OPS) calls, often showing a very short downsweep at the start and steep droop at the end. At a distance, the initial downsweep and terminal droop may not be detected, leaving a call which looks almost linear, and dropping to about 16 kHz.
May show regular frequency alternation, with the higher pulses being more curved, steeper, and dropping to about 18 kHz with a minimum slope of about 10 OPS. No feeding buzz recorded, but nature of frequency alternation and attack-phase suggest a Molosside rather than an Emballonuride, since the higher pulses are very different in shape to the lower pulses. High pulses may be irregularly interspersed amongst lower pulses.
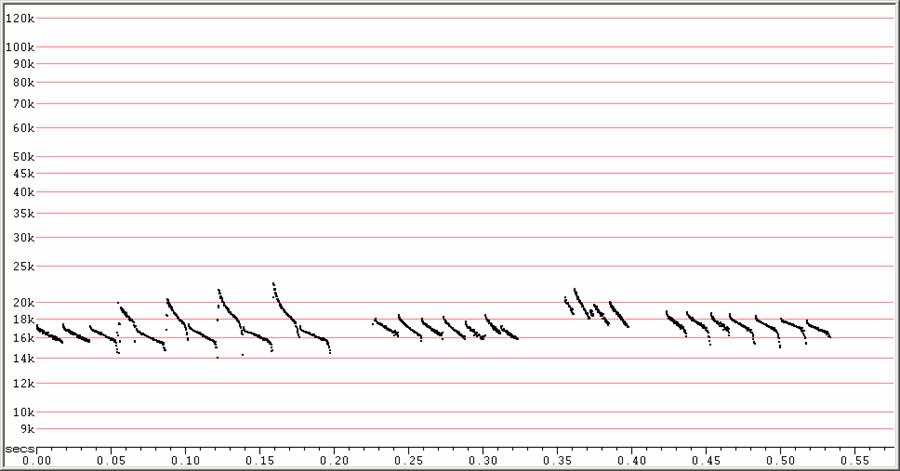
F6, Divratio 8, compressed.
Several search-phase sequences showing frequency alternation at left
.
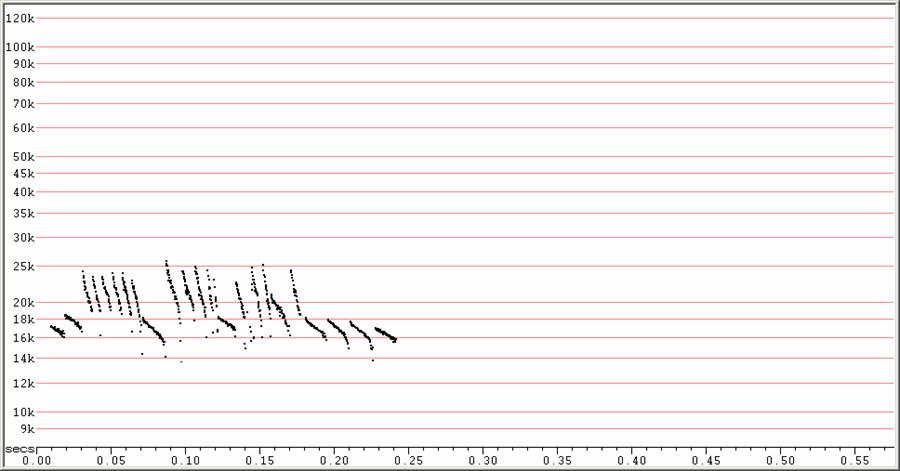
F6, Divratio 8, compressed.
A single sequence all from the same bat. The steeper calls in this
sequence were
produced at a much higher repetition rate than the lower, flatter calls,
and
therefore appear to represent attack-phase calls. It is likely the bat was
just
too far away to detect the feeding buzz.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home
None of these was recorded more than a very few times, but they show a variety of call shapes and frequencies with unknown relationships to the call types described above. They are presented here as further examples of calls which have not been associated with any species.
The calls depicted in B, below, look very similar to calls attributed to Cheiromeles torquatus (see above). Those in A would most likely represent calls in very low clutter from either Cheiromeles torquatus or Mops mops, assuming these are the only Molosside contenders in the region.
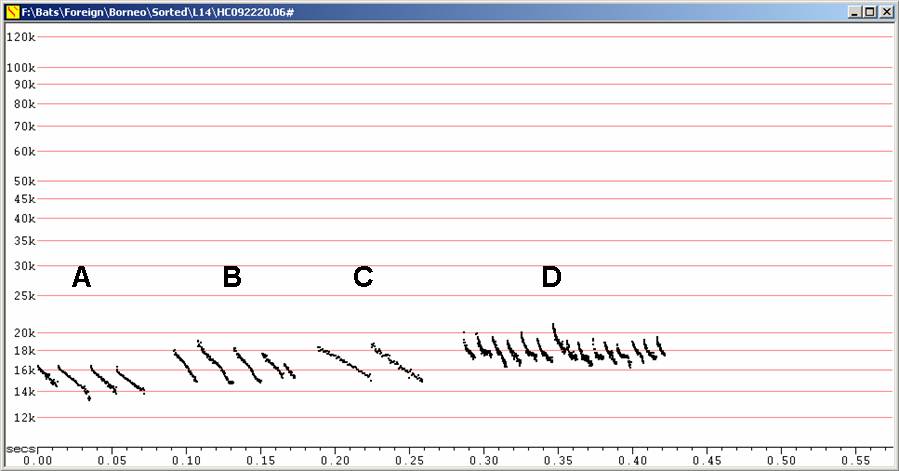
Various low frequency calls at F6, compressed.
Back to: Index of call types, top of page
Back to: Bat Notes contents, Anabat Contents, Home